
The endoplasmic reticulum (ER) is a part of a transportation system of the eukaryotic cell, and has many other important functions such as protein folding. It is a type of organelle made up of two subunits – rough endoplasmic reticulum (RER), and smooth endoplasmic reticulum (SER). The endoplasmic reticulum is found in most eukaryotic cells and forms an interconnected network of flattened, membrane-enclosed sacs known as cisternae, and tubular structures in the SER. The membranes of the ER are continuous with the outer nuclear membrane. The endoplasmic reticulum is not found in red blood cells, or spermatozoa.
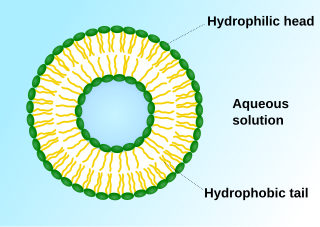
In cell biology, a vesicle is a structure within or outside a cell, consisting of liquid or cytoplasm enclosed by a lipid bilayer. Vesicles form naturally during the processes of secretion (exocytosis), uptake (endocytosis), and the transport of materials within the plasma membrane. Alternatively, they may be prepared artificially, in which case they are called liposomes. If there is only one phospholipid bilayer, the vesicles are called unilamellar liposomes; otherwise they are called multilamellar liposomes. The membrane enclosing the vesicle is also a lamellar phase, similar to that of the plasma membrane, and intracellular vesicles can fuse with the plasma membrane to release their contents outside the cell. Vesicles can also fuse with other organelles within the cell. A vesicle released from the cell is known as an extracellular vesicle.

The lipid bilayer is a thin polar membrane made of two layers of lipid molecules. These membranes are flat sheets that form a continuous barrier around all cells. The cell membranes of almost all organisms and many viruses are made of a lipid bilayer, as are the nuclear membrane surrounding the cell nucleus, and membranes of the membrane-bound organelles in the cell. The lipid bilayer is the barrier that keeps ions, proteins and other molecules where they are needed and prevents them from diffusing into areas where they should not be. Lipid bilayers are ideally suited to this role, even though they are only a few nanometers in width, because they are impermeable to most water-soluble (hydrophilic) molecules. Bilayers are particularly impermeable to ions, which allows cells to regulate salt concentrations and pH by transporting ions across their membranes using proteins called ion pumps.
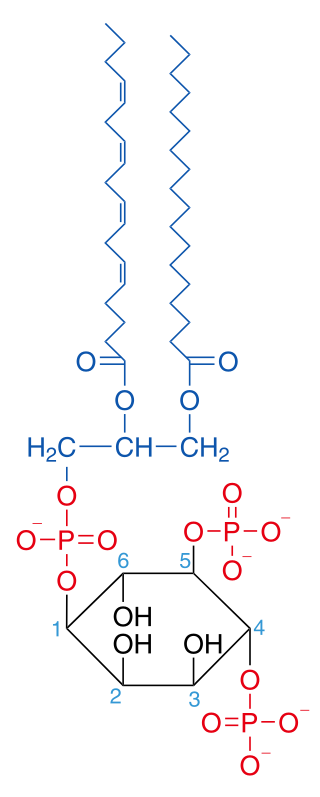
Phosphatidylinositol 4,5-bisphosphate or PtdIns(4,5)P2, also known simply as PIP2 or PI(4,5)P2, is a minor phospholipid component of cell membranes. PtdIns(4,5)P2 is enriched at the plasma membrane where it is a substrate for a number of important signaling proteins. PIP2 also forms lipid clusters that sort proteins.

Scramblase is a protein responsible for the translocation of phospholipids between the two monolayers of a lipid bilayer of a cell membrane. In humans, phospholipid scramblases (PLSCRs) constitute a family of five homologous proteins that are named as hPLSCR1–hPLSCR5. Scramblases are members of the general family of transmembrane lipid transporters known as flippases. Scramblases are distinct from flippases and floppases. Scramblases, flippases, and floppases are three different types of enzymatic groups of phospholipid transportation enzymes. The inner-leaflet, facing the inside of the cell, contains negatively charged amino-phospholipids and phosphatidylethanolamine. The outer-leaflet, facing the outside environment, contains phosphatidylcholine and sphingomyelin. Scramblase is an enzyme, present in the cell membrane, that can transport (scramble) the negatively charged phospholipids from the inner-leaflet to the outer-leaflet, and vice versa.

ADP ribosylation factors (ARFs) are members of the ARF family of GTP-binding proteins of the Ras superfamily. ARF family proteins are ubiquitous in eukaryotic cells, and six highly conserved members of the family have been identified in mammalian cells. Although ARFs are soluble, they generally associate with membranes because of N-terminus myristoylation. They function as regulators of vesicular traffic and actin remodelling.
In molecular biology, an annexin A5 affinity assay is a test to quantify the number of cells undergoing apoptosis. The assay uses the protein annexin A5 to tag apoptotic and dead cells, and the numbers are then counted using either flow cytometry or a fluorescence microscope.

Annexin A2 also known as annexin II is a protein that in humans is encoded by the ANXA2 gene.

Annexin A5 is a cellular protein in the annexin group. In flow cytometry, annexin V is commonly used to detect apoptotic cells by its ability to bind to phosphatidylserine, a marker of apoptosis when it is on the outer leaflet of the plasma membrane. The function of the protein is unknown; however, annexin A5 has been proposed to play a role in the inhibition of blood coagulation by competing for phosphatidylserine binding sites with prothrombin and also to inhibit the activity of phospholipase A1. These properties have been found by in vitro experiments.

Ezrin also known as cytovillin or villin-2 is a protein that in humans is encoded by the EZR gene.

Annexin A6 is a protein that in humans is encoded by the ANXA6 gene.

S100 calcium-binding protein A10 (S100A10), also known as p11, is a protein that is encoded by the S100A10 gene in humans and the S100a10 gene in other species. S100A10 is a member of the S100 family of proteins containing two EF-hand calcium-binding motifs. S100 proteins are localized in the cytoplasm and/or nucleus of a wide range of cells. They regulate a number of cellular processes such as cell cycle progression and differentiation. The S100 protein is implicated in exocytosis and endocytosis by reorganization of F-actin.

S100 calcium-binding protein A11 (S100A11) is a protein that in humans is encoded by the S100A11 gene.

Annexin A4 is a protein that in humans is encoded by the ANXA4 gene.

Annexin A7 is a protein that in humans is encoded by the ANXA7 gene.
Annexin A8-like protein 2 is a protein that in humans is encoded by the ANXA8L2 gene.
Scinderin is a protein that in humans is encoded by the SCIN gene. Scinderin is an actin severing protein belonging to the gelsolin superfamily. It was discovered in Dr. Trifaro's laboratory at the University of Ottawa, Canada. Secretory tissues are rich in scinderin. In these tissues scinderin, a calcium dependent protein, regulates cortical actin networks. Normally secretory vesicles are excluded from release sites on the plasma membrane by the presence of a cortical actin filament network. During cell stimulation, calcium channels open allowing calcium ions to enter the secretory cell. Increase in intracellular calcium activates scinderin with the consequent actin filament severing and local dissociation of actin filament networks. This allows the movement of secretory vesicles to release sites on the plasma membrane.
Annexin A9 is a protein that in humans is encoded by the ANXA9 gene.
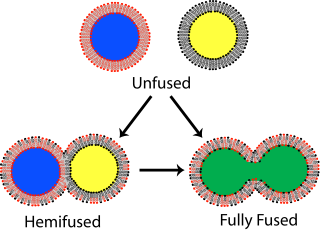
In membrane biology, fusion is the process by which two initially distinct lipid bilayers merge their hydrophobic cores, resulting in one interconnected structure. If this fusion proceeds completely through both leaflets of both bilayers, an aqueous bridge is formed and the internal contents of the two structures can mix. Alternatively, if only one leaflet from each bilayer is involved in the fusion process, the bilayers are said to be hemifused. In hemifusion, the lipid constituents of the outer leaflet of the two bilayers can mix, but the inner leaflets remain distinct. The aqueous contents enclosed by each bilayer also remain separated.

Ferlins are an ancient protein family involved in vesicle fusion and membrane trafficking. Ferlins are distinguished by their multiple tandem C2 domains, and sometimes a FerA and a DysF domain. Mutations in ferlins can cause human diseases such as muscular dystrophy and deafness. Abnormalities in expression of myoferlin, a human ferlin protein, is also directly associated with higher mortality rate and tumor recurrence in several types of cancer, including pancreatic, colorectal, breast, cervical, stomach, ovarian, cervical, thyroid, endometrial, and oropharyngeal squamous cell carcinoma. In other animals, ferlin mutations can cause infertility.
