Related Research Articles
Calcium channel blockers (CCB), calcium channel antagonists or calcium antagonists are a group of medications that disrupt the movement of calcium through calcium channels. Calcium channel blockers are used as antihypertensive drugs, i.e., as medications to decrease blood pressure in patients with hypertension. CCBs are particularly effective against large vessel stiffness, one of the common causes of elevated systolic blood pressure in elderly patients. Calcium channel blockers are also frequently used to alter heart rate, to prevent peripheral and cerebral vasospasm, and to reduce chest pain caused by angina pectoris.
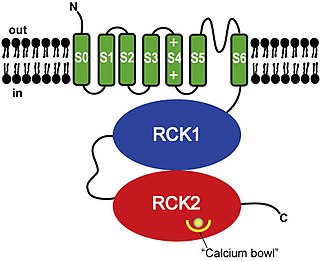
BK channels (big potassium), are large conductance calcium-activated potassium channels, also known as Maxi-K, slo1, or Kca1.1. BK channels are voltage-gated potassium channels that conduct large amounts of potassium ions (K+) across the cell membrane, hence their name, big potassium. These channels can be activated (opened) by either electrical means, or by increasing Ca2+ concentrations in the cell. BK channels help regulate physiological processes, such as circadian behavioral rhythms and neuronal excitability. BK channels are also involved in many processes in the body, as it is a ubiquitous channel. They have a tetrameric structure that is composed of a transmembrane domain, voltage sensing domain, potassium channel domain, and a cytoplasmic C-terminal domain, with many X-ray structures for reference. Their function is to repolarize the membrane potential by allowing for potassium to flow outward, in response to a depolarization or increase in calcium levels.
An inhibitory postsynaptic potential (IPSP) is a kind of synaptic potential that makes a postsynaptic neuron less likely to generate an action potential. The opposite of an inhibitory postsynaptic potential is an excitatory postsynaptic potential (EPSP), which is a synaptic potential that makes a postsynaptic neuron more likely to generate an action potential. IPSPs can take place at all chemical synapses, which use the secretion of neurotransmitters to create cell-to-cell signalling. EPSPs and IPSPs compete with each other at numerous synapses of a neuron. This determines whether an action potential occurring at the presynaptic terminal produces an action potential at the postsynaptic membrane. Some common neurotransmitters involved in IPSPs are GABA and glycine.

Voltage-gated ion channels are a class of transmembrane proteins that form ion channels that are activated by changes in a cell's electrical membrane potential near the channel. The membrane potential alters the conformation of the channel proteins, regulating their opening and closing. Cell membranes are generally impermeable to ions, thus they must diffuse through the membrane through transmembrane protein channels.
A calcium channel is an ion channel which shows selective permeability to calcium ions. It is sometimes synonymous with voltage-gated calcium channel, which are a type of calcium channel regulated by changes in membrane potential. Some calcium channels are regulated by the binding of a ligand. Other calcium channels can also be regulated by both voltage and ligands to provide precise control over ion flow. Some cation channels allow calcium as well as other cations to pass through the membrane.
Voltage-gated calcium channels (VGCCs), also known as voltage-dependent calcium channels (VDCCs), are a group of voltage-gated ion channels found in the membrane of excitable cells (e.g., muscle, glial cells, neurons, etc.) with a permeability to the calcium ion Ca2+. These channels are slightly permeable to sodium ions, so they are also called Ca2+–Na+ channels, but their permeability to calcium is about 1000-fold greater than to sodium under normal physiological conditions.
Molecular neuroscience is a branch of neuroscience that observes concepts in molecular biology applied to the nervous systems of animals. The scope of this subject covers topics such as molecular neuroanatomy, mechanisms of molecular signaling in the nervous system, the effects of genetics and epigenetics on neuronal development, and the molecular basis for neuroplasticity and neurodegenerative diseases. As with molecular biology, molecular neuroscience is a relatively new field that is considerably dynamic.

Sodium channels are integral membrane proteins that form ion channels, conducting sodium ions (Na+) through a cell's membrane. They belong to the superfamily of cation channels.
Sodium voltage-gated channel alpha subunit 9 is a sodium ion channel that in humans is encoded by the SCN9A gene. It is usually expressed at high levels in two types of neurons: the nociceptive (pain) neurons at the dorsal root ganglion (DRG) and trigeminal ganglion; and sympathetic ganglion neurons, which are part of the autonomic (involuntary) nervous system.
The P-type calcium channel is a type of voltage-dependent calcium channel. Similar to many other high-voltage-gated calcium channels, the α1 subunit determines most of the channel's properties. The 'P' signifies cerebellar Purkinje cells, referring to the channel's initial site of discovery. P-type calcium channels play a similar role to the N-type calcium channel in neurotransmitter release at the presynaptic terminal and in neuronal integration in many neuronal types.

The L-type calcium channel is part of the high-voltage activated family of voltage-dependent calcium channel. "L" stands for long-lasting referring to the length of activation. This channel has four isoforms: Cav1.1, Cav1.2, Cav1.3, and Cav1.4.

Sodium channel, voltage-gated, type XI, alpha subunit also known as SCN11A or Nav1.9 is a voltage-gated sodium ion channel protein which is encoded by the SCN11A gene on chromosome 3 in humans. Like Nav1.7 and Nav1.8, Nav1.9 plays a role in pain perception. This channel is largely expressed in small-diameter nociceptors of the dorsal root ganglion and trigeminal ganglion neurons, but is also found in intrinsic myenteric neurons.

Potassium voltage-gated channel, Shab-related subfamily, member 1, also known as KCNB1 or Kv2.1, is a protein that, in humans, is encoded by the KCNB1 gene.

Calcium channel, voltage-dependent, L type, alpha 1D subunit is a protein that in humans is encoded by the CACNA1D gene. Cav1.3 channels belong to the Cav1 family, which form L-type calcium currents and are sensitive to selective inhibition by dihydropyridines (DHP).
Low-threshold spikes (LTS) refer to membrane depolarizations by the T-type calcium channel. LTS occur at low, negative, membrane depolarizations. They often follow a membrane hyperpolarization, which can be the result of decreased excitability or increased inhibition. LTS result in the neuron reaching the threshold for an action potential. LTS is a large depolarization due to an increase in Ca2+ conductance, so LTS is mediated by calcium (Ca2+) conductance. The spike is typically crowned by a burst of two to seven action potentials, which is known as a low-threshold burst. LTS are voltage dependent and are inactivated if the cell's resting membrane potential is more depolarized than −60mV. LTS are deinactivated, or recover from inactivation, if the cell is hyperpolarized and can be activated by depolarizing inputs, such as excitatory postsynaptic potentials (EPSP). LTS were discovered by Rodolfo Llinás and coworkers in the 1980s.
Hyperpolarization-activated cyclic nucleotide–gated (HCN) channels are integral membrane proteins that serve as nonselective voltage-gated cation channels in the plasma membranes of heart and brain cells. HCN channels are sometimes referred to as pacemaker channels because they help to generate rhythmic activity within groups of heart and brain cells. HCN channels are activated by membrane hyperpolarization, are permeable to Na + and K +, and are constitutively open at voltages near the resting membrane potential. HCN channels are encoded by four genes and are widely expressed throughout the heart and the central nervous system.
Calcium channel, voltage-dependent, T type, alpha 1H subunit, also known as CACNA1H, is a protein which in humans is encoded by the CACNA1H gene.

Spike-and-wave is a pattern of the electroencephalogram (EEG) typically observed during epileptic seizures. A spike-and-wave discharge is a regular, symmetrical, generalized EEG pattern seen particularly during absence epilepsy, also known as ‘petit mal’ epilepsy. The basic mechanisms underlying these patterns are complex and involve part of the cerebral cortex, the thalamocortical network, and intrinsic neuronal mechanisms.

In neurophysiology, a dendritic spike refers to an action potential generated in the dendrite of a neuron. Dendrites are branched extensions of a neuron. They receive electrical signals emitted from projecting neurons and transfer these signals to the cell body, or soma. Dendritic signaling has traditionally been viewed as a passive mode of electrical signaling. Unlike its axon counterpart which can generate signals through action potentials, dendrites were believed to only have the ability to propagate electrical signals by physical means: changes in conductance, length, cross sectional area, etc. However, the existence of dendritic spikes was proposed and demonstrated by W. Alden Spencer, Eric Kandel, Rodolfo Llinás and coworkers in the 1960s and a large body of evidence now makes it clear that dendrites are active neuronal structures. Dendrites contain voltage-gated ion channels giving them the ability to generate action potentials. Dendritic spikes have been recorded in numerous types of neurons in the brain and are thought to have great implications in neuronal communication, memory, and learning. They are one of the major factors in long-term potentiation.
Calcium channel, voltage-dependent, T type, alpha 1G subunit, also known as CACNA1G or Cav3.1 is a protein which in humans is encoded by the CACNA1G gene. It is one of the primary targets in the pharmacology of absence seizure.
References
- 1 2 3 4 5 6 7 8 Catterall WA (August 2011). "Voltage-gated calcium channels". Cold Spring Harbor Perspectives in Biology. 3 (8): a003947. doi:10.1101/cshperspect.a003947. PMC 3140680 . PMID 21746798.
- 1 2 Iftinca MC (May 2011). "Neuronal T-type calcium channels: what's new? Iftinca: T-type channel regulation". Journal of Medicine and Life. 4 (2): 126–138. PMC 3124264 . PMID 21776294.
- 1 2 3 4 5 6 Perez-Reyes E (January 2003). "Molecular physiology of low-voltage-activated t-type calcium channels". Physiological Reviews. 83 (1): 117–161. doi:10.1152/physrev.00018.2002. PMID 12506128.
- 1 2 3 4 5 6 7 Dziegielewska B, Gray LS, Dziegielewski J (April 2014). "T-type calcium channels blockers as new tools in cancer therapies". Pflügers Archiv. 466 (4): 801–810. doi:10.1007/s00424-014-1444-z. PMID 24449277. S2CID 18238236.
- 1 2 3 4 5 Nelson MT, Todorovic SM, Perez-Reyes E (2006). "The role of T-type calcium channels in epilepsy and pain". Current Pharmaceutical Design. 12 (18): 2189–2197. doi:10.2174/138161206777585184. PMID 16787249.
- 1 2 3 4 Todorovic SM, Jevtovic-Todorovic V (April 2014). "Targeting of CaV3.2 T-type calcium channels in peripheral sensory neurons for the treatment of painful diabetic neuropathy". Pflügers Archiv. 466 (4): 701–706. doi:10.1007/s00424-014-1452-z. PMID 24482063. S2CID 15152953.
- 1 2 Bermejo PE, Anciones B (September 2009). "A review of the use of zonisamide in Parkinson's disease". Therapeutic Advances in Neurological Disorders. 2 (5): 313–317. doi:10.1177/1756285609338501. PMC 3002602 . PMID 21180621.
- 1 2 Nicholls JG, Martin AR, Fuchs PA, Brown DA, Diamond ME, Weisblat DA (2012). From neuron to brain (5th ed.). Sunderland, Mass.: Sinauer Associates. pp. 87–88. ISBN 9780878936090.
- ↑ Kessler SK, McGinnis E (February 2019). "A Practical Guide to Treatment of Childhood Absence Epilepsy". Paediatric Drugs. 21 (1): 15–24. doi:10.1007/s40272-019-00325-x. PMC 6394437 . PMID 30734897.
- ↑ Choi S, Na HS, Kim J, Lee J, Lee S, Kim D, et al. (July 2007). "Attenuated pain responses in mice lacking Ca(V)3.2 T-type channels". Genes, Brain and Behavior. 6 (5): 425–431. doi: 10.1111/j.1601-183X.2006.00268.x . PMID 16939637. S2CID 21984943.
- ↑ Bourinet E, Francois A, Laffray S (February 2016). "T-type calcium channels in neuropathic pain" (PDF). Pain. 157 (Suppl 1): S15–S22. doi:10.1097/j.pain.0000000000000469. PMID 26785151. S2CID 23847088.
- ↑ Kerckhove N, Mallet C, François A, Boudes M, Chemin J, Voets T, et al. (April 2014). "Ca(v)3.2 calcium channels: the key protagonist in the supraspinal effect of paracetamol" (PDF). Pain. 155 (4): 764–772. doi:10.1016/j.pain.2014.01.015. PMID 24447516. S2CID 5241999.
- ↑ Marger F, Gelot A, Alloui A, Matricon J, Ferrer JF, Barrère C, et al. (July 2011). "T-type calcium channels contribute to colonic hypersensitivity in a rat model of irritable bowel syndrome". Proceedings of the National Academy of Sciences of the United States of America. 108 (27): 11268–11273. Bibcode:2011PNAS..10811268M. doi: 10.1073/pnas.1100869108 . PMC 3131334 . PMID 21690417.
- ↑ Rubin JE, McIntyre CC, Turner RS, Wichmann T (July 2012). "Basal ganglia activity patterns in parkinsonism and computational modeling of their downstream effects". The European Journal of Neuroscience. 36 (2): 2213–2228. doi:10.1111/j.1460-9568.2012.08108.x. PMC 3400124 . PMID 22805066.
- 1 2 Devergnas A, Chen E, Ma Y, Hamada I, Pittard D, Kammermeier S, et al. (January 2016). "Anatomical localization of Cav3.1 calcium channels and electrophysiological effects of T-type calcium channel blockade in the motor thalamus of MPTP-treated monkeys". Journal of Neurophysiology. 115 (1): 470–485. doi:10.1152/jn.00858.2015. PMC 4760490 . PMID 26538609.
- ↑ Albin RL, Young AB, Penney JB (October 1989). "The functional anatomy of basal ganglia disorders". Trends in Neurosciences. 12 (10): 366–375. doi:10.1016/0166-2236(89)90074-X. hdl: 2027.42/28186 . PMID 2479133. S2CID 8112392.
- ↑ Bosch-Bouju C, Hyland BI, Parr-Brownlie LC (2013-01-01). "Motor thalamus integration of cortical, cerebellar and basal ganglia information: implications for normal and parkinsonian conditions". Frontiers in Computational Neuroscience. 7: 163. doi: 10.3389/fncom.2013.00163 . PMC 3822295 . PMID 24273509.
- ↑ Leventhal DK, Gage GJ, Schmidt R, Pettibone JR, Case AC, Berke JD (February 2012). "Basal ganglia beta oscillations accompany cue utilization". Neuron. 73 (3): 523–536. doi:10.1016/j.neuron.2011.11.032. PMC 3463873 . PMID 22325204.
- ↑ Xiang Z, Thompson AD, Brogan JT, Schulte ML, Melancon BJ, Mi D, et al. (December 2011). "The Discovery and Characterization of ML218: A Novel, Centrally Active T-Type Calcium Channel Inhibitor with Robust Effects in STN Neurons and in a Rodent Model of Parkinson's Disease". ACS Chemical Neuroscience. 2 (12): 730–742. doi:10.1021/cn200090z. PMC 3285241 . PMID 22368764.