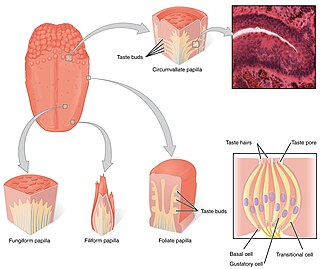
Localization of G14 and G-gustducin in type II taste cells. TrpM5 is a cation channel of the TRP superfamily that is highly expressed in taste buds of the tongue. In vallate and foliate taste buds of transgenic mice expressing TrpM5-GFP (green), two G-protein alpha subunits, G14 (red) and G-gustducin (blue), exist within the population of type II taste cells that express TrpM5. The red G14 staining that is present in sweet-sensitive taste cells outlines the reactive cells, showing that it is membrane-associated
Gustducin is a G protein associated with taste and the gustatory system, found in some taste receptor cells. Research on the discovery and isolation of gustducin is recent. It is known to play a large role in the transduction of bitter, sweet and umami stimuli. Its pathways (especially for detecting bitter stimuli) are many and diverse.
An intriguing feature of gustducin is its similarity to transducin. These two G proteins have been shown to be structurally and functionally similar, leading researchers to believe that the sense of taste evolved in a similar fashion to the sense of sight.
Gustducin is a heterotrimeric protein composed of the products of the GNAT3 (α-subunit), GNB1 (β-subunit) and GNG13 (γ-subunit).
Discovery
Gustducin was discovered in 1992 when degenerate oligonucleotide primers were synthesized and mixed with a taste tissue cDNA library. The DNA products were amplified by the polymerase chain reaction method, and eight positive clones were shown to encode the α subunits of G-proteins, (which interact with G-protein-coupled receptors). Of these eight, two had previously been shown to encode rod and cone α-transducin. The eighth clone, α-gustducin, was unique to the gustatory tissue.[1]
Comparisons with transducin
Upon analyzing the amino-acid sequence of α-gustducin, it was discovered that α-gustducins and α-transducins were closely related. This work showed that α-gustducin's protein sequence gives it 80% identity to both rod and cone a-transducin. Despite the structural similarities, the two proteins have very different functionalities.
However, the two proteins have similar mechanism and capabilities. Transducin removes the inhibition from cGMPPhosphodiesterase, which leads to the breakdown of cGMP. Similarly, α-gustducin binds the inhibitory subunits of taste cell cAMP Phosphodiesterase which causes a decrease in cAMP levels. Also, the terminal 38 amino acids of α-gustducin and α-transducin are identical. This suggests that gustducin can interact with opsin and opsin-like G-coupled receptors. Conversely, this also suggests that transducin can interact with taste receptors.
The structural similarities between gustducin and transducin are so great that comparison with transducin were used to propose a model of gustducin's role and functionality in taste transduction.[citation needed]
Other G protein α-subunits have been identified in TRCs (e.g. Gαi-2, Gαi-3, Gα14, Gα15, Gαq, Gαs) with function that has not yet been determined.[2]
Location
While gustducin was known to be expressed in some taste receptor cells (TRCs), studies with rats showed that gustducin was also present in a limited subset of cells lining the stomach and intestine. These cells appear to share several feature of TRCs. Another study with humans brought to light two immunoreactive patterns for α-gustducin in human circumavallate and foliate taste cells: plasmalemmal and cytosolic. These two studies showed that gustducin is distributed through gustatory tissue and some gastric and intestinal tissue and gustducin is presented either in the cytoplasm or in apical membranes on TRC surfaces.
Research showed that bitter-stimulated type 2 taste receptors (T2R/TRB) are only found in taste receptor cells positive for the expression of gustducin. α-Gustducin is selectively expressed in ~25–30% of TRCs [2]
Evolution of the gustducin-mediated signaling model
Due to its structural similarity to transducin, gustducin was predicted to activate a phosphodiesterase (PDE). Phosphodieterases were found in taste tissues and their activation was tested in vitro with both gustducin and transducin. This experiment revealed transducin and gustducin were both expressed in taste tissue (1:25 ratio) and that both G proteins are capable of activating retinal PDE. Furthermore, when present with denatonium and quinine, both G proteins can activate taste specific PDEs. This indicated that both gustducin and transducin are important in the signal transduction of denatonium and quinine.
The 1992 research also investigated the role of gustducin in bitter taste reception by using "knock-out" mice lacking the gene for α-gustducin. A taste test with knock-out and control mice revealed that the knock-out mice showed no preference between bitter and regular food in most cases. When the α-gustducin gene was re-inserted into the knock-out mice, the original taste ability returned.
However, the loss of the α-gustducin gene did not completely remove the ability of the knock-out mice to taste bitter food, indicating that α-gustducin is not the only mechanism for tasting bitter food. It was thought at the time that an alternative mechanism of bitter taste detection could be associated with the βγ subunit of gustducin. This theory was later validated when it was discovered that both peripheral and central gustatory neurons typically respond to more than one type of taste stimulant, although a neuron typically would favor one specific stimulant over others. This suggests that, while many neurons favor bitter taste stimuli, neurons that favor other stimuli such as sweet and umami may be capable of detecting bitter stimuli in the absence of bitter stimulant receptors, as with the knock-out mice.[citation needed]
Second messengers IP3 and cAMP
Until recently, the nature of gustducin and its second messengers was unclear. It was clear, however, that gustducin transduced intracellular signals. Spielman was one of the first to look at the speed of taste reception, utilizing the quenched-flow technique. When the taste cells were exposed to the bitter stimulants denatonium and sucrose octaacetate, the intracellular response - a transient increase of IP3 - occurred within 50-100 millisecond of stimulation. This was not unexpected, as it was known that transducin was capable of sending signals within rod and cone cells at similar speeds. This indicated that IP3 was one of the second messengers used in bitter taste transduction. It was later discovered that cAMP also causes an influx of cations during bitter and some sweet taste transduction, leading to the conclusion that it also acted as a second messenger to gustducin.[citation needed]
Bitter transduction
When bitter-stimulated T2R/TRB receptors activate gustducin heterotrimers, gustducin acts to mediate two responses in taste receptor cells: a decrease in cAMPs triggered by α-gustducin, and a rise in IP3(Inositol trisphosphate) and diacylglycerol (DAG) from βγ-gustducin.[2]
Although the following steps of the α-gustducin pathway are unconfirmed, it is suspected that decreased cAMPs may act on protein kinases which would regulate taste receptor cell ion channel activity. It is also possible that cNMP levels directly regulate the activity of cNMP-gated channels and cNMP-inhibited ion channels expressed in taste receptor cells. The βγ-gustducin pathway continues with the activation of IP3 receptors and the release of Ca2+ followed by neurotransmitter release.[citation needed]
Bitter taste transduction models Several models have been suggested for the mechanisms regarding the transduction of bitter taste signals.
Cell-surface receptors: Patch clamping experiments have shown evidence that bitter compounds such as denatonium and sucrose octaacetate act directly on specific cell-surface receptors.[citation needed]
Direct activation of G proteins: Certain bitter stimulants such as quinine have been shown to activate G proteins directly. While these mechanisms have been identified,[by whom?] the physiologic relevance of the mechanism has not yet been established.
PDE activation: Other bitter compounds, such as thioacetamide and propylthiouracil, have been shown[by whom?] to have stimulatory effects on PDEs. This mechanism has been recognized in bovine tongue epithelium contains fungiform papillae.
PDE inhibition: Other bitter compounds have been shown[by whom?] to inhibit PDE. Bacitracin and hydrochloride have been shown to inhibit PDE in bovine taste tissue
Channel blockage: Patch clamping experiments have shown that several bitter ions act directly on potassium channels, blocking them. This suggests that the potassium channels would be located in the apical region of the taste cells. While this theory seems[by whom?] valid, it has only been identified in mudpuppy taste cells.
It is thought[by whom?] that these five diverse mechanisms have developed as defense mechanisms. This would imply that many different poisonous or harmful bitter agents exist and these five mechanisms exist to prevent humans from eating or drinking them. It is also possible that some mechanisms can act as backups should a primary mechanism fail. One example of this could be quinine, which has been shown to both inhibit and activate PDE in bovine taste tissue.
Sweet transduction
There are currently two models proposed for sweet taste transduction. The first pathway is a GPCRGs-cAMP pathway. This pathway starts with sucrose and other sugars activating Gs inside the cell through a membrane-bound GPCR. The activated Gas activates adenylyl cyclase to generate cAMP. From this point, one of two pathways can be taken. cAMP may act directly to cause an influx of cations through cAMP- gated channels or cAMP can activate protein kinase A, which causes the phosphorylation of K+ channels, thus closing the channels, allowing for depolarization of the taste cell, subsequent opening of voltage-gated Ca2+ channels and causing neurotransmitter release [citation needed].
The second pathway is a GPCR-Gq/Gβγ-IP3 pathway which is used with artificial sweeteners. Artificial sweeteners bind and activate GPCRs coupled to PLCβ2 by either α-Gq or Gβγ. The activated subunits activate PLCβ2 to generate IP3 and DAG. IP3 and DAG elicit Ca2+ release from the endoplasmic reticulum and cause cellular depolarization. An influx of Ca2+ triggers neurotransmitter release. While these two pathways coexist in the same TRCs, it is unclear how the receptors selectively mediate cAMP responses to sugars and IP3 responses to artificial sweeteners [citation needed].
Evolution of bitter taste receptors
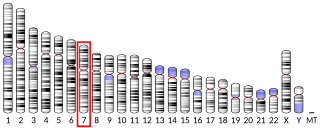
Of the five basic tastes, three (sweet, bitter and umami tastes) are mediated by receptors from the G protein-coupled receptor family. Mammalian bitter taste receptors (T2Rs) are encoded by a gene family of only a few dozen members. It is believed that bitter taste receptors evolved as a mechanism to avoid ingesting poisonous and harmful substances. If this is the case, one might expect different species to develop different bitter taste receptors based on dietary and geographical constraints. With the exception of T2R1 (which lies on chromosome 5) all human bitter taste receptor genes can be found clustered on chromosome 7 and chromosome 12. Analyzing the relationships between bitter taste receptor genes show that the genes on the same chromosome are more closely related to each other than genes on different chromosomes. Furthermore, the genes on chromosome 12 have higher sequence similarity than the genes found on chromosome 7. This indicates that these genes evolved via tandem gene duplications and that chromosome 12, as a result of its higher sequence similarity between its genes, went through these tandem duplications more recently than the genes on chromosome 7.
Gustducin in the stomach
Recent work by Enrique Rozengurt has shed some light on the presence of gustducin in the stomach and gastrointestinal tract.[3] His work suggests that gustducin is present in these areas as a defense mechanism. It is widely known that some drugs and toxins can cause harm and even be lethal if ingested. It has already been theorized that multiple bitter taste reception pathways exist to prevent harmful substances from being ingested, but a person can choose to ignore the taste of a substance. Ronzegurt suggests that the presence of gustducin in epithelial cells in the stomach and gastrointestinal tract are indicative of another line of defense against ingested toxins. Whereas taste cells in the mouth are designed to compel a person to spit out a toxin, these stomach cells may act to force a person to spit up the toxins in the form of vomit.
G protein-coupled receptors (GPCRs), also known as seven-(pass)-transmembrane domain receptors, 7TM receptors, heptahelical receptors, serpentine receptors, and G protein-linked receptors (GPLR), form a large group of evolutionarily related proteins that are cell surface receptors that detect molecules outside the cell and activate cellular responses. They are coupled with G proteins. They pass through the cell membrane seven times in the form of six loops of amino acid residues, which is why they are sometimes referred to as seven-transmembrane receptors. Ligands can bind either to the extracellular N-terminus and loops or to the binding site within transmembrane helices. They are all activated by agonists, although a spontaneous auto-activation of an empty receptor has also been observed.
G proteins, also known as guanine nucleotide-binding proteins, are a family of proteins that act as molecular switches inside cells, and are involved in transmitting signals from a variety of stimuli outside a cell to its interior. Their activity is regulated by factors that control their ability to bind to and hydrolyze guanosine triphosphate (GTP) to guanosine diphosphate (GDP). When they are bound to GTP, they are 'on', and, when they are bound to GDP, they are 'off'. G proteins belong to the larger group of enzymes called GTPases.
Second messengers are intracellular signaling molecules released by the cell in response to exposure to extracellular signaling molecules—the first messengers. Second messengers trigger physiological changes at cellular level such as proliferation, differentiation, migration, survival, apoptosis and depolarization.
3′,5′-cyclic-nucleotide phosphodiesterases (EC 3.1.4.17) are a family of phosphodiesterases. Generally, these enzymes hydrolyze a nucleoside 3′,5′-cyclic phosphate to a nucleoside 5′-phosphate:
A taste receptor or tastant is a type of cellular receptor which facilitates the sensation of taste. When food or other substances enter the mouth, molecules interact with saliva and are bound to taste receptors in the oral cavity and other locations. Molecules which give a sensation of taste are considered "sapid".
Gi protein alpha subunit is a family of heterotrimeric G protein alpha subunits. This family is also commonly called the Gi/o family or Gi/o/z/t family to include closely related family members. G alpha subunits may be referred to as Gi alpha, Gαi, or Giα.
Taste receptor type 2 member 3 is a protein that in humans is encoded by the TAS2R3 gene.
Taste receptor type 2 member 4 is a protein that in humans is encoded by the TAS2R4 gene.
Taste receptor type 2 member 8 is a protein that in humans is encoded by the TAS2R8 gene.
Taste receptor type 2 member 9 is a protein that in humans is encoded by the TAS2R9 gene.
Taste receptor type 2 member 14 is a protein that in humans is encoded by the TAS2R14 gene.
T1R2 - Taste receptor type 1 member 2 is a protein that in humans is encoded by the TAS1R2 gene.
Taste receptor type 1 member 3 is a protein that in humans is encoded by the TAS1R3 gene. The TAS1R3 gene encodes the human homolog of mouse Sac taste receptor, a major determinant of differences between sweet-sensitive and -insensitive mouse strains in their responsiveness to sucrose, saccharin, and other sweeteners.
—
Guanine nucleotide-binding protein subunit beta-4 is a protein that in humans is encoded by the GNB4 gene.
Guanine nucleotide-binding protein G(I)/G(S)/G(O) subunit gamma-13 is a protein that in humans is encoded by the GNG13 gene.
Solitary chemosensory cells (SCCs) are isolated elements located in epithelia of the apparatuses of endodermic origin. In the aquatic vertebrates, SCCs are also present in the skin. In oral cavity, SCCs precedes the development of taste buds. For long time, SCCs were considered to be typical of aquatic vertebrates. Recently, these elements were also demonstrated in mammals.
In the field of molecular biology, the cAMP-dependent pathway, also known as the adenylyl cyclase pathway, is a G protein-coupled receptor-triggered signaling cascade used in cell communication.
Guanine nucleotide-binding protein G(t) subunit alpha-3, also known as gustducin alpha-3 chain, is a protein subunit that in humans is encoded by the GNAT3 gene.
Robert F. Margolskee is an American academic. He is the director of the Monell Chemical Senses Center and adjunct professor in the Department of Neuroscience at the Mount Sinai School of Medicine. Margolskee is also the a co-founder of Redpoint Bio. Margolskee has been a pioneer in the application of molecular biology and transgenic animal models to the study of taste transduction and chemosensation. He has made numerous seminal discoveries in the taste field, including the identification and molecular cloning of taste specific receptors, G proteins, channels and other taste signal transduction elements.
↑ Rozengurt E (August 2006). "Taste receptors in the gastrointestinal tract. I. Bitter taste receptors and alpha-gustducin in the mammalian gut". Am. J. Physiol. Gastrointest. Liver Physiol. 291 (2): G171–7. doi:10.1152/ajpgi.00073.2006. PMID16710053.
Lindemann B (April 1999). "Receptor seeks ligand: on the way to cloning the molecular receptors for sweet and bitter taste". Nat. Med. 5 (4): 381–2. doi:10.1038/7377. PMID10202923. S2CID5650076.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.