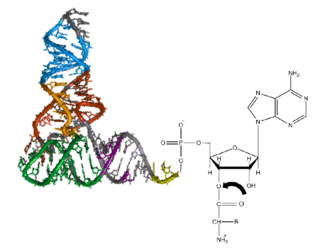An aminoacyl-tRNA synthetase (aaRS or ARS), also called tRNA-ligase, is an enzyme that attaches the appropriate amino acid onto its corresponding tRNA. It does so by catalyzing the transesterification of a specific cognate amino acid or its precursor to one of all its compatible cognate tRNAs to form an aminoacyl-tRNA. In humans, the 20 different types of aa-tRNA are made by the 20 different aminoacyl-tRNA synthetases, one for each amino acid of the genetic code.
This is sometimes called "charging" or "loading" the tRNA with an amino acid. Once the tRNA is charged, a ribosome can transfer the amino acid from the tRNA onto a growing peptide, according to the genetic code. Aminoacyl tRNA therefore plays an important role in RNA translation, the expression of genes to create proteins.
Mechanism
The synthetase first binds ATP and the corresponding amino acid (or its precursor) to form an aminoacyl-adenylate, releasing inorganic pyrophosphate (PPi). The adenylate-aaRS complex then binds the appropriate tRNA molecule's D arm, and the amino acid is transferred from the aa-AMP to either the 2'- or the 3'-OH of the last tRNA nucleotide (A76) at the 3'-end.
The mechanism can be summarized in the following reaction series:
Summing the reactions, the highly exergonic overall reaction is as follows:
Amino Acid + tRNA + ATP → Aminoacyl-tRNA + AMP + PPi
Some synthetases also mediate an editing reaction to ensure high fidelity of tRNA charging. If the incorrect tRNA is added (aka. the tRNA is found to be improperly charged), the aminoacyl-tRNA bond is hydrolyzed. This can happen when two amino acids have different properties even if they have similar shapes—as is the case with valine and threonine.
The accuracy of aminoacyl-tRNA synthetase is so high that it is often paired with the word "superspecificity” when it is compared to other enzymes that are involved in metabolism. Although not all synthetases have a domain with the sole purpose of editing, they make up for it by having specific binding and activation of their affiliated amino acids. Another contribution to the accuracy of these synthetases is the ratio of concentrations of aminoacyl-tRNA synthetase and its cognate tRNA. Since tRNA synthetase improperly acylates the tRNA when the synthetase is overproduced, a limit must exist on the levels of aaRSs and tRNAs in vivo.[1][2]
Classes
There are two classes of aminoacyl tRNA synthetase, each composed of ten enzymes:[3][4]
Class II has three highly conserved sequence motifs. It aminoacylates at the 3'-OH of a terminal adenosine on tRNA, and is usually dimeric or tetrameric (two or four subunits, respectively). Although phenylalanine-tRNA synthetase is class II, it aminoacylates at the 2'-OH.
The amino acids are attached to the hydroxyl (-OH) group of the adenosine via the carboxyl (-COOH) group.
Regardless of where the aminoacyl is initially attached to the nucleotide, the 2'-O-aminoacyl-tRNA will ultimately migrate to the 3' position via transesterification.
Bacterial aminoacyl-tRNA synthetases can be grouped as follows:[5]
Class
Amino acids
I
Arg, Cys, Gln, Glu, Ile, Leu, Met, Trp, Tyr, Val
II
Ala, Asn, Asp, Gly, His, Lys, Pro, Phe, Ser, Thr
Amino acids which use class II aaRS seem to be evolutionarily older.[6]
A general structure of an aminoacyl-tRNA synthetase is shown here with an editing site as well as an activation site. The main difference between class I and class II synthetases is the activation site. Here you can see the general structure of the Rossmann fold seen in class I aaRSs and the general structure of antiparallel beta-sheets seen in class II aaRSs.Alignment of the core domains of aminoacyl-tRNA synthetases class I and class II. Essential binding site residues (Backbone Brackets and Arginine Tweezers) are colored. N-terminal residues are highlighted in blue, C-terminal in red.
Structures
Both classes of aminoacyl-tRNA synthetases are multidomain proteins. In a typical scenario, an aaRS consists of a catalytic domain (where both the above reactions take place) and an anticodon binding domain (which interacts mostly with the anticodon region of the tRNA). Transfer-RNAs for different amino acids differ not only in their anticodon but also at other points, giving them slightly different overall configurations. The aminoacyl-tRNA synthetases recognize the correct tRNAs primarily through their overall configuration, not just through their anticodon.[7] In addition, some aaRSs have additional RNA binding domains and editing domains[8] that cleave incorrectly paired aminoacyl-tRNA molecules.
The catalytic domains of all the aaRSs of a given class are found to be homologous to one another, whereas class I and class II aaRSs are unrelated to one another. The class I aaRSs feature a cytidylyltransferase-like Rossmann fold seen in proteins like glycerol-3-phosphate cytidylyltransferase, nicotinamide nucleotide adenylyltransferase and archaeal FAD synthase, whereas the class II aaRSs have a unique fold related to biotin and lipoate ligases.
Aminoacyl-tRNA synthetases have been kinetically studied, showing that Mg2+ ions play an active catalytic role and therefore aaRs have a degree of magnesium dependence. Increasing the Mg2+ concentration leads to an increase in the equilibrium constants for the aminoacyl-tRNA synthetases’ reactions. Although this trend was seen in both class I and class II synthetases, the magnesium dependence for the two classes are very distinct. Class II synthetases have two or (more frequently) three Mg2+ ions, while class I only requires one Mg2+ ion.[10][11]
Beside their lack of overall sequence and structure similarity, class I and class II synthetases feature different ATP recognition mechanisms. While class I binds via interactions mediated by backbone hydrogen bonds, class II uses a pair of arginine residues to establish salt bridges to its ATP ligand. This oppositional implementation is manifested in two structural motifs, the Backbone Brackets and Arginine Tweezers, which are observable in all class I and class II structures, respectively. The high structural conservation of these motifs suggest that they must have been present since ancient times.[12]
Evolution
Most of the aaRSs of a given specificity are evolutionarily closer to one another than to aaRSs of another specificity. However, AsnRS and GlnRS group within AspRS and GluRS, respectively. Most of the aaRSs of a given specificity also belong to a single class. However, there are two distinct versions of the LysRS - one belonging to the class I family and the other belonging to the class II family.[citation needed]
The molecular phylogenies of aaRSs are often not consistent with accepted organismal phylogenies. That is, they violate the so-called canonical phylogenetic pattern shown by most other enzymes for the three domains of life - Archaea, Bacteria, and Eukarya. Furthermore, the phylogenies inferred for aaRSs of different amino acids often do not agree with one another. In addition, aaRS paralogs within the same species show a high degree of divergence between them. These are clear indications that horizontal transfer has occurred several times during the evolutionary history of aaRSs.[13]
A widespread belief in the evolutionary stability of this superfamily, meaning that every organism has all the aaRSs for their corresponding amino acids, is misconceived. A large-scale genomic analysis on ~2500 prokaryotic genomes showed that many of them miss one or more aaRS genes whereas many genomes have 1 or more paralogs.[14] AlaRS, GlyRS, LeuRS, IleRS and ValRS are the most evolutionarily stable members of the family. GluRS, LysRS and CysRS often have paralogs, whereas AsnRS, GlnRS, PylRS and SepRS are often absent from many genomes.
With the exception of AlaRS, it has been discovered that 19 out of the 20 human aaRSs have added at least one new domain or motif.[15] These new domains and motifs vary in function and are observed in various forms of life. A common novel function within human aaRSs is providing additional regulation of biological processes. There exists a theory that the increasing number of aaRSs that add domains is due to the continuous evolution of higher organisms with more complex and efficient building blocks and biological mechanisms. One key piece of evidence to this theory is that after a new domain is added to an aaRS, the domain becomes fully integrated. This new domain's functionality is conserved from that point on.[16]
As genetic efficiency evolved in higher organisms, 13 new domains with no obvious association with the catalytic activity of aaRSs genes have been added.
Application in biotechnology
In some of the aminoacyl tRNA synthetases, the cavity that holds the amino acid can be mutated and modified to carry unnatural amino acids synthesized in the lab, and to attach them to specific tRNAs. This expands the genetic code, beyond the twenty canonical amino acids found in nature, to include an unnatural amino acid as well. The unnatural amino acid is coded by a nonsense (TAG, TGA, TAA) triplet, a quadruplet codon, or in some cases a redundant rare codon. The organism that expresses the mutant synthetase can then be genetically programmed to incorporate the unnatural amino acid into any desired position in any protein of interest, allowing biochemists or structural biologists to probe or change the protein's function. For instance, one can start with the gene for a protein that binds a certain sequence of DNA, and, by directing an unnatural amino acid with a reactive side-chain into the binding site, create a new protein that cuts the DNA at the target-sequence, rather than binding it.
By mutating aminoacyl tRNA synthetases, chemists have expanded the genetic codes of various organisms to include lab-synthesized amino acids with all kinds of useful properties: photoreactive, metal-chelating, xenon-chelating, crosslinking, spin-resonant, fluorescent, biotinylated, and redox-active amino acids.[17] Another use is introducing amino acids bearing reactive functional groups for chemically modifying the target protein.
Certain diseases’ causation (such as neuronal pathologies, cancer, disturbed metabolic conditions, and autoimmune disorders) have been correlated to specific mutations of aminoacyl-tRNA synthetases. Charcot-Marie-Tooth (CMT) is the most frequent heritable disorder of the peripheral nervous system (a neuronal disease) and is caused by a heritable mutation in glycol-tRNA and tyrosyl-tRNA.[18] Diabetes, a metabolic disease, induces oxidative stress, which triggers a build up of mitochondrial tRNA mutations. It has also been discovered that tRNA synthetases may be partially involved in the etiology of cancer.[19] A high level of expression or modification of aaRSs has been observed within a range of cancers. A common outcome from mutations of aaRSs is a disturbance of dimer shape/formation which has a direct relationship with its function. These correlations between aaRSs and certain diseases have opened up a new door to synthesizing therapeutics.[20]
Noncatalytic domains
The novel domain additions to aaRS genes are accretive and progressive up the Tree of Life.[21][22][23] The strong evolutionary pressure for these small non-catalytic protein domains suggested their importance.[24] Findings beginning in 1999 and later revealed a previously unrecognized layer of biology: these proteins control gene expression within the cell of origin, and when released exert homeostatic and developmental control in specific human cell types, tissues and organs during adult or fetal development or both, including pathways associated with angiogenesis, inflammation, the immune response, the mechanistic target of rapamycin (mTOR) signalling, apoptosis, tumorigenesis, and interferon gamma (IFN-γ) and p53 signalling.[25][26][27][28][29][30][31][32][33]
Substrate Depletion
In 2022, it was discovered that aminoacyl-trna synthetases may incorporate alternative amino acids during shortages of their precursors. In particular, tryptophanyl-tRNA synthetase (WARS1) will incorporate phenylalanine during tryptophan depletion, essentially inducing a W>F codon reassignment.[34] Depletion of the other substrate of aminoacyl-tRNA synthetases, the cognate tRNA, may be relevant to certain diseases, e.g. Charcot–Marie–Tooth disease. It was shown that CMT-mutant glycyl-tRNA synthetase variants are still able to bind tRNA-gly but fail to release it, leading to depletion of the cellular pool of glycyl-tRNA-gly, what in turn results in stalling of the ribosome on glycine codons during mRNA translation.[35]
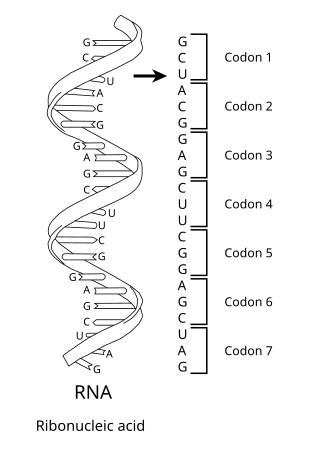
The genetic code is the set of rules used by living cells to translate information encoded within genetic material into proteins. Translation is accomplished by the ribosome, which links proteinogenic amino acids in an order specified by messenger RNA (mRNA), using transfer RNA (tRNA) molecules to carry amino acids and to read the mRNA three nucleotides at a time. The genetic code is highly similar among all organisms and can be expressed in a simple table with 64 entries.
Pyrrolysine is an α-amino acid that is used in the biosynthesis of proteins in some methanogenic archaea and bacteria; it is not present in humans. It contains an α-amino group, a carboxylic acid group. Its pyrroline side-chain is similar to that of lysine in being basic and positively charged at neutral pH.
In biology, translation is the process in living cells in which proteins are produced using RNA molecules as templates. The generated protein is a sequence of amino acids. This sequence is determined by the sequence of nucleotides in the RNA. The nucleotides are considered three at a time. Each such triple results in addition of one specific amino acid to the protein being generated. The matching from nucleotide triple to amino acid is called the genetic code. The translation is performed by a large complex of functional RNA and proteins called ribosomes. The entire process is called gene expression.
Transfer RNA is an adaptor molecule composed of RNA, typically 76 to 90 nucleotides in length, that serves as the physical link between the mRNA and the amino acid sequence of proteins. Transfer RNA (tRNA) does this by carrying an amino acid to the protein-synthesizing machinery of a cell called the ribosome. Complementation of a 3-nucleotide codon in a messenger RNA (mRNA) by a 3-nucleotide anticodon of the tRNA results in protein synthesis based on the mRNA code. As such, tRNAs are a necessary component of translation, the biological synthesis of new proteins in accordance with the genetic code.
A wobble base pair is a pairing between two nucleotides in RNA molecules that does not follow Watson-Crick base pair rules. The four main wobble base pairs are guanine-uracil (G-U), hypoxanthine-uracil (I-U), hypoxanthine-adenine (I-A), and hypoxanthine-cytosine (I-C). In order to maintain consistency of nucleic acid nomenclature, "I" is used for hypoxanthine because hypoxanthine is the nucleobase of inosine; nomenclature otherwise follows the names of nucleobases and their corresponding nucleosides. The thermodynamic stability of a wobble base pair is comparable to that of a Watson-Crick base pair. Wobble base pairs are fundamental in RNA secondary structure and are critical for the proper translation of the genetic code.
Aminoacyl-tRNA is tRNA to which its cognate amino acid is chemically bonded (charged). The aa-tRNA, along with particular elongation factors, deliver the amino acid to the ribosome for incorporation into the polypeptide chain that is being produced during translation.
EF-Tu is a prokaryotic elongation factor responsible for catalyzing the binding of an aminoacyl-tRNA (aa-tRNA) to the ribosome. It is a G-protein, and facilitates the selection and binding of an aa-tRNA to the A-site of the ribosome. As a reflection of its crucial role in translation, EF-Tu is one of the most abundant and highly conserved proteins in prokaryotes. It is found in eukaryotic mitochondria as TUFM.
Glycine—tRNA ligase also known as glycyl–tRNA synthetase is an enzyme that in humans is encoded by the GARS1 gene.
In enzymology, a phenylalanine—tRNA ligase is an enzyme that catalyzes the chemical reaction
In enzymology, a threonine-tRNA ligase is an enzyme that catalyzes the chemical reaction
Tyrosine—tRNA ligase, also known as tyrosyl-tRNA synthetase is an enzyme that is encoded by the gene YARS. Tyrosine—tRNA ligase catalyzes the chemical reaction
Tyrosyl-tRNA synthetase, cytoplasmic, also known as Tyrosine-tRNA ligase, is an enzyme that in humans is encoded by the YARS gene.
Glutaminyl-tRNA synthetase is an enzyme that in humans is encoded by the QARS gene.
Phenylalanyl-tRNA synthetase alpha chain is an enzyme that in humans is encoded by the FARSA gene.
An expanded genetic code is an artificially modified genetic code in which one or more specific codons have been re-allocated to encode an amino acid that is not among the 22 common naturally-encoded proteinogenic amino acids.
Amino acid activation refers to the attachment of an amino acid to its respective transfer RNA (tRNA). The reaction occurs in the cell cytosol and consists of two steps: first, the enzyme aminoacyl tRNA synthetase catalyzes the binding of adenosine triphosphate (ATP) to a corresponding amino acid, forming a reactive aminoacyl adenylate intermediate and releasing inorganic pyrophosphate (PPi). Subsequently, aminoacyl tRNA synthetase binds the AMP-amino acid to a tRNA molecule, releasing AMP and attaching the amino acid to the tRNA. The resulting aminoacyl-tRNA is said to be charged.
Aminoacyl-tRNA synthetases, class II is a family of proteins. These proteins catalyse the attachment of an amino acid to its cognate transfer RNA molecule in a highly specific two-step reaction. These proteins differ widely in size and oligomeric state, and have a limited sequence homology.
The aminoacyl-tRNA synthetases catalyse the attachment of an amino acid to its cognate transfer RNA molecule in a highly specific two-step reaction. These proteins differ widely in size and oligomeric state, and have limited sequence homology. The 20 aminoacyl-tRNA synthetases are divided into two classes, I and II. Class I aminoacyl-tRNA synthetases contain a characteristic Rossmann fold catalytic domain and are mostly monomeric. Class II aminoacyl-tRNA synthetases share an anti-parallel beta-sheet fold flanked by alpha-helices, and are mostly dimeric or multimeric, containing at least three conserved regions. However, tRNA binding involves an alpha-helical structure that is conserved between class I and class II synthetases. In reactions catalysed by the class I aminoacyl-tRNA synthetases, the aminoacyl group is coupled to the 2'-hydroxyl of the tRNA, while, in class II reactions, the 3'-hydroxyl site is preferred. The synthetases specific for arginine, cysteine, glutamic acid, glutamine, isoleucine, leucine, methionine, tyrosine, tryptophan and valine belong to class I synthetases; these synthetases are further divided into three subclasses, a, b and c, according to sequence homology. The synthetases specific for alanine, asparagine, aspartic acid, glycine, histidine, lysine, phenylalanine, proline, serine, and threonine belong to class-II synthetases.
Xiang-Lei Yang (杨湘磊) is a Chinese-born American molecular biologist. She is a professor at The Scripps Research Institute, located in La Jolla, California. Her work has contributed to the establishment of physiological importance of aminoacyl-tRNA synthetases beyond their classical role in supporting mRNA translation and their disordered processes that contribute to disease. She founded the Translation Machinery in Health and Disease Gordon Research Conference, an ongoing biannual international conference since 2015. She helped co-found aTyr Pharma, a Nasdaq-listed biotechnology company.
Cyclodipeptide synthases (CDPSs) are a newly defined family of peptide-bond forming enzymes that are responsible for the ribosome-independent biosynthesis of various cyclodipeptides, which are the precursors of many natural products with important biological activities. As a substrate for this synthesis, CDPSs use two amino acids activated as aminoacyl-tRNAs (aa-tRNAs), therefore diverting them from the ribosomal machinery. The first member of this family was identified in 2002 during the characterization of the albonoursin biosynthetic pathway in Streptomyces noursei. CDPSs are present in bacteria, fungi, and animal cells.
References
↑ McClain WH (November 1993). "Rules that govern tRNA identity in protein synthesis". Journal of Molecular Biology. 234 (2): 257–80. doi:10.1006/jmbi.1993.1582. PMID8230212.
↑ Swanson R, Hoben P, Sumner-Smith M, Uemura H, Watson L, Söll D (December 1988). "Accuracy of in vivo aminoacylation requires proper balance of tRNA and aminoacyl-tRNA synthetase". Science. 242 (4885): 1548–51. Bibcode:1988Sci...242.1548S. doi:10.1126/science.3144042. PMID3144042.
↑ Lareau LF, Green RE, Bhatnagar RS, Brenner SE (June 2004). "The evolving roles of alternative splicing". Current Opinion in Structural Biology. 14 (3): 273–82. doi:10.1016/j.sbi.2004.05.002. PMID15193306.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.