Related Research Articles
Ribonuclease is a type of nuclease that catalyzes the degradation of RNA into smaller components. Ribonucleases can be divided into endoribonucleases and exoribonucleases, and comprise several sub-classes within the EC 2.7 and 3.1 classes of enzymes.
Ribonuclease H is a family of non-sequence-specific endonuclease enzymes that catalyze the cleavage of RNA in an RNA/DNA substrate via a hydrolytic mechanism. Members of the RNase H family can be found in nearly all organisms, from bacteria to archaea to eukaryotes.

Transfer RNA is an adaptor molecule composed of RNA, typically 76 to 90 nucleotides in length, that serves as the physical link between the mRNA and the amino acid sequence of proteins. Transfer RNA (tRNA) does this by carrying an amino acid to the protein synthesizing machinery of a cell called the ribosome. Complementation of a 3-nucleotide codon in a messenger RNA (mRNA) by a 3-nucleotide anticodon of the tRNA results in protein synthesis based on the mRNA code. As such, tRNAs are a necessary component of translation, the biological synthesis of new proteins in accordance with the genetic code.
Polyadenylation is the addition of a poly(A) tail to an RNA transcript, typically a messenger RNA (mRNA). The poly(A) tail consists of multiple adenosine monophosphates; in other words, it is a stretch of RNA that has only adenine bases. In eukaryotes, polyadenylation is part of the process that produces mature mRNA for translation. In many bacteria, the poly(A) tail promotes degradation of the mRNA. It, therefore, forms part of the larger process of gene expression.
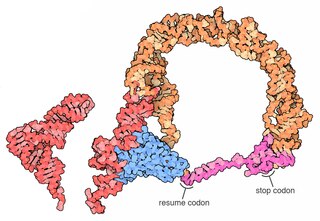
Transfer-messenger RNA is a bacterial RNA molecule with dual tRNA-like and messenger RNA-like properties. The tmRNA forms a ribonucleoprotein complex (tmRNP) together with Small Protein B (SmpB), Elongation Factor Tu (EF-Tu), and ribosomal protein S1. In trans-translation, tmRNA and its associated proteins bind to bacterial ribosomes which have stalled in the middle of protein biosynthesis, for example when reaching the end of a messenger RNA which has lost its stop codon. The tmRNA is remarkably versatile: it recycles the stalled ribosome, adds a proteolysis-inducing tag to the unfinished polypeptide, and facilitates the degradation of the aberrant messenger RNA. In the majority of bacteria these functions are carried out by standard one-piece tmRNAs. In other bacterial species, a permuted ssrA gene produces a two-piece tmRNA in which two separate RNA chains are joined by base-pairing.
Ribonuclease P is a type of ribonuclease which cleaves RNA. RNase P is unique from other RNases in that it is a ribozyme – a ribonucleic acid that acts as a catalyst in the same way that a protein-based enzyme would. Its function is to cleave off an extra, or precursor, sequence of RNA on tRNA molecules. Further, RNase P is one of two known multiple turnover ribozymes in nature, the discovery of which earned Sidney Altman and Thomas Cech the Nobel Prize in Chemistry in 1989: in the 1970s, Altman discovered the existence of precursor tRNA with flanking sequences and was the first to characterize RNase P and its activity in processing of the 5' leader sequence of precursor tRNA. Recent findings also reveal that RNase P has a new function. It has been shown that human nuclear RNase P is required for the normal and efficient transcription of various small noncoding RNAs, such as tRNA, 5S rRNA, SRP RNA and U6 snRNA genes, which are transcribed by RNA polymerase III, one of three major nuclear RNA polymerases in human cells.
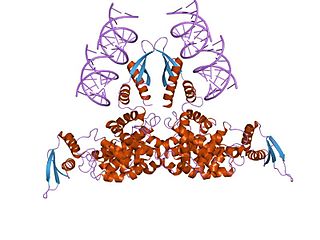
Ribonuclease III (RNase III or RNase C)(BRENDA 3.1.26.3) is a type of ribonuclease that recognizes dsRNA and cleaves it at specific targeted locations to transform them into mature RNAs. These enzymes are a group of endoribonucleases that are characterized by their ribonuclease domain, which is labelled the RNase III domain. They are ubiquitous compounds in the cell and play a major role in pathways such as RNA precursor synthesis, RNA Silencing, and the pnp autoregulatory mechanism.
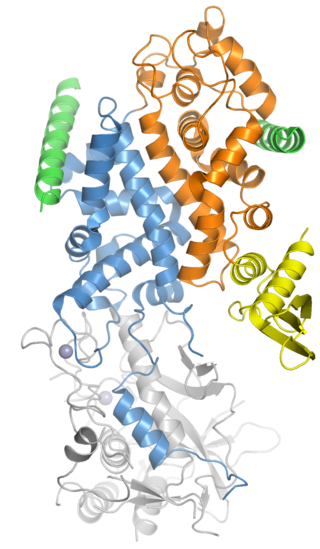
Drosha is a Class 2 ribonuclease III enzyme that in humans is encoded by the DROSHA gene. It is the primary nuclease that executes the initiation step of miRNA processing in the nucleus. It works closely with DGCR8 and in correlation with Dicer. It has been found significant in clinical knowledge for cancer prognosis and HIV-1 replication.
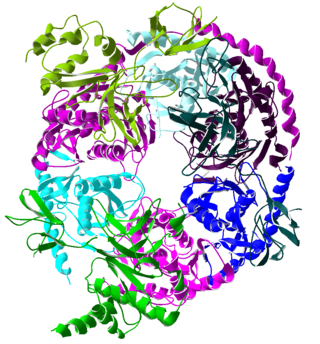
The exosome complex is a multi-protein intracellular complex capable of degrading various types of RNA molecules. Exosome complexes are found in both eukaryotic cells and archaea, while in bacteria a simpler complex called the degradosome carries out similar functions.
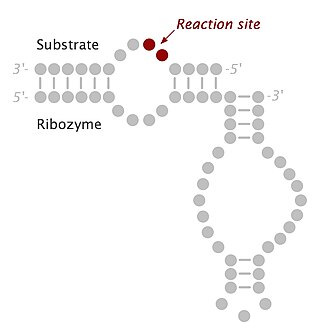
The hairpin ribozyme is a small section of RNA that can act as a ribozyme. Like the hammerhead ribozyme it is found in RNA satellites of plant viruses. It was first identified in the minus strand of the tobacco ringspot virus (TRSV) satellite RNA where it catalyzes self-cleavage and joining (ligation) reactions to process the products of rolling circle virus replication into linear and circular satellite RNA molecules. The hairpin ribozyme is similar to the hammerhead ribozyme in that it does not require a metal ion for the reaction.
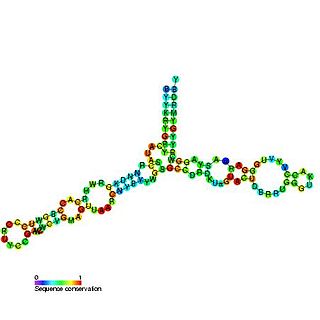
The 5S ribosomal RNA is an approximately 120 nucleotide-long ribosomal RNA molecule with a mass of 40 kDa. It is a structural and functional component of the large subunit of the ribosome in all domains of life, with the exception of mitochondrial ribosomes of fungi and animals. The designation 5S refers to the molecule's sedimentation velocity in an ultracentrifuge, which is measured in Svedberg units (S).
Exoribonuclease II is an enzyme. This enzyme catalyses the following chemical reaction

Ribonuclease pancreatic is an enzyme that in humans is encoded by the RNASE1 gene.
Zinc phosphodiesterase ELAC protein 2 is an enzyme that in humans is encoded by the ELAC2 gene. on chromosome 17. It is an endonuclease thought to be involved in mitochondrial tRNA maturation,

Zinc phosphodiesterase ELAC protein 1 is an enzyme that in humans is encoded by the ELAC1 gene.
Ribonuclease E is a bacterial ribonuclease that participates in the processing of ribosomal RNA and the chemical degradation of bulk cellular RNA.

The retroviral ribonuclease H is a catalytic domain of the retroviral reverse transcriptase (RT) enzyme. The RT enzyme is used to generate complementary DNA (cDNA) from the retroviral RNA genome. This process is called reverse transcription. To complete this complex process, the retroviral RT enzymes need to adopt a multifunctional nature. They therefore possess 3 of the following biochemical activities: RNA-dependent DNA polymerase, ribonuclease H, and DNA-dependent DNA polymerase activities. Like all RNase H enzymes, the retroviral RNase H domain cleaves DNA/RNA duplexes and will not degrade DNA or unhybridized RNA.
tRNA-intron lyase is an enzyme. As an endonuclease enzyme, tRNA-intron lyase is responsible for splicing phosphodiester bonds within non-coding ribonucleic acid chains. These non-coding RNA molecules form tRNA molecules after being processed, and this is dependent on tRNA-intron lyase to splice the pretRNA. tRNA processing is an important post-transcriptional modification necessary for tRNA maturation because it locates and removes introns in the pretRNA. This enzyme catalyses the following chemical reaction:

Ribonuclease V1 is a ribonuclease enzyme found in the venom of the Caspian cobra. It cleaves double-stranded RNA in a non-sequence-specific manner, usually requiring a substrate of at least six stacked nucleotides. Like many ribonucleases, the enzyme requires the presence of magnesium ions for activity.
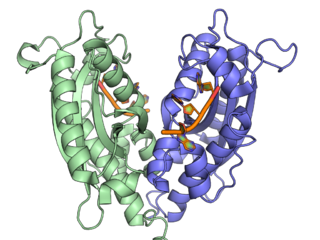
Ribonuclease T is a ribonuclease enzyme involved in the maturation of transfer RNA and ribosomal RNA in bacteria, as well as in DNA repair pathways. It is a member of the DnaQ family of exonucleases and non-processively acts on the 3' end of single-stranded nucleic acids. RNase T is capable of cleaving both DNA and RNA, with extreme sequence specificity discriminating against cytosine at the 3' end of the substrate.
References
- ↑ Schiffer S, Rösch S, Marchfelder A (June 2002). "Assigning a function to a conserved group of proteins: the tRNA 3'-processing enzymes". The EMBO Journal. 21 (11): 2769–77. doi:10.1093/emboj/21.11.2769. PMC 126033 . PMID 12032089.
- ↑ Mayer M, Schiffer S, Marchfelder A (February 2000). "tRNA 3' processing in plants: nuclear and mitochondrial activities differ". Biochemistry. 39 (8): 2096–105. doi:10.1021/bi992253e. PMID 10684660.
- ↑ Schiffer S, Helm M, Théobald-Dietrich A, Giegé R, Marchfelder A (July 2001). "The plant tRNA 3' processing enzyme has a broad substrate spectrum". Biochemistry. 40 (28): 8264–72. doi:10.1021/bi0101953. PMID 11444972.
- ↑ Kunzmann A, Brennicke A, Marchfelder A (January 1998). "5' end maturation and RNA editing have to precede tRNA 3' processing in plant mitochondria". Proceedings of the National Academy of Sciences of the United States of America. 95 (1): 108–13. doi: 10.1073/pnas.95.1.108 . PMC 18142 . PMID 9419337.
- ↑ Mörl M, Marchfelder A (January 2001). "The final cut. The importance of tRNA 3'-processing". EMBO Reports. 2 (1): 17–20. doi:10.1093/embo-reports/kve006. PMC 1083803 . PMID 11252717.
- ↑ Minagawa A, Takaku H, Takagi M, Nashimoto M (April 2004). "A novel endonucleolytic mechanism to generate the CCA 3' termini of tRNA molecules in Thermotoga maritima". The Journal of Biological Chemistry. 279 (15): 15688–97. doi: 10.1074/jbc.M313951200 . PMID 14749326.
- ↑ Takaku H, Minagawa A, Takagi M, Nashimoto M (May 2003). "A candidate prostate cancer susceptibility gene encodes tRNA 3' processing endoribonuclease". Nucleic Acids Research. 31 (9): 2272–8. doi:10.1093/nar/gkg337. PMC 154223 . PMID 12711671.
- ↑ Wilusz JE, Freier SM, Spector DL (November 2008). "3' end processing of a long nuclear-retained noncoding RNA yields a tRNA-like cytoplasmic RNA". Cell. 135 (5): 919–32. doi:10.1016/j.cell.2008.10.012. PMC 2722846 . PMID 19041754.