
The cochlea is the part of the inner ear involved in hearing. It is a spiral-shaped cavity in the bony labyrinth, in humans making 2.75 turns around its axis, the modiolus. A core component of the cochlea is the organ of Corti, the sensory organ of hearing, which is distributed along the partition separating the fluid chambers in the coiled tapered tube of the cochlea.

The basilar membrane is a stiff structural element within the cochlea of the inner ear which separates two liquid-filled tubes that run along the coil of the cochlea, the scala media and the scala tympani. The basilar membrane moves up and down in response to incoming sound waves, which are converted to traveling waves on the basilar membrane.
The organ of Corti, or spiral organ, is the receptor organ for hearing and is located in the mammalian cochlea. This highly varied strip of epithelial cells allows for transduction of auditory signals into nerve impulses' action potential. Transduction occurs through vibrations of structures in the inner ear causing displacement of cochlear fluid and movement of hair cells at the organ of Corti to produce electrochemical signals.

The auditory system is the sensory system for the sense of hearing. It includes both the sensory organs and the auditory parts of the sensory system.
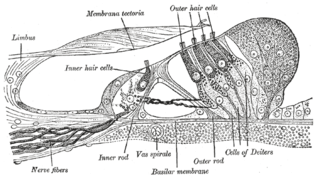
Hair cells are the sensory receptors of both the auditory system and the vestibular system in the ears of all vertebrates, and in the lateral line organ of fishes. Through mechanotransduction, hair cells detect movement in their environment.

Chloride channels are a superfamily of poorly understood ion channels specific for chloride. These channels may conduct many different ions, but are named for chloride because its concentration in vivo is much higher than other anions. Several families of voltage-gated channels and ligand-gated channels have been characterized in humans.

In the inner ear, stereocilia are the mechanosensing organelles of hair cells, which respond to fluid motion in numerous types of animals for various functions, including hearing and balance. They are about 10–50 micrometers in length and share some similar features of microvilli. The hair cells turn the fluid pressure and other mechanical stimuli into electric stimuli via the many microvilli that make up stereocilia rods. Stereocilia exist in the auditory and vestibular systems.
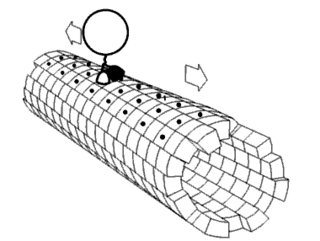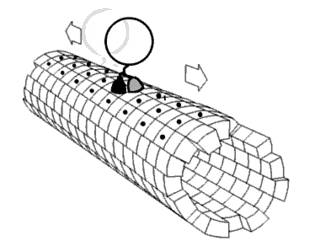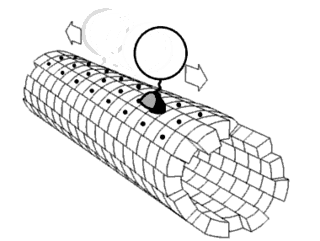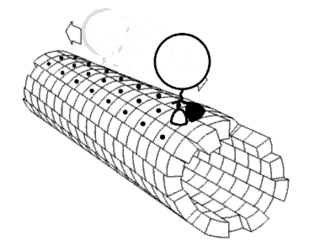
Motor proteins are a class of molecular motors that can move along the cytoplasm of cells. They convert chemical energy into mechanical work by the hydrolysis of ATP. Flagellar rotation, however, is powered by a proton pump.
The tectoria membrane (TM) is one of two acellular membranes in the cochlea of the inner ear, the other being the basilar membrane (BM). "Tectorial" in anatomy means forming a cover. The TM is located above the spiral limbus and the spiral organ of Corti and extends along the longitudinal length of the cochlea parallel to the BM. Radially the TM is divided into three zones, the limbal, middle and marginal zones. Of these the limbal zone is the thinnest (transversally) and overlies the auditory teeth of Huschke with its inside edge attached to the spiral limbus. The marginal zone is the thickest (transversally) and is divided from the middle zone by Hensen's Stripe. It overlies the sensory inner hair cells and electrically-motile outer hair cells of the organ of Corti and during acoustic stimulation stimulates the inner hair cells through fluid coupling, and the outer hair cells via direct connection to their tallest stereocilia.
Pendrin is an anion exchange protein that in humans is encoded by the SLC26A4 gene . Pendrin was initially identified as a sodium-independent chloride-iodide exchanger with subsequent studies showing that it also accepts formate and bicarbonate as substrates. Pendrin is similar to the Band 3 transport protein found in red blood cells. Pendrin is the protein which is mutated in Pendred syndrome, which is an autosomal recessive disorder characterized by sensorineural hearing loss, goiter and a partial organification problem detectable by a positive perchlorate test.

Voltage-dependent anion channels, or mitochondrial porins, are a class of porin ion channel located on the outer mitochondrial membrane. There is debate as to whether or not this channel is expressed in the cell surface membrane.
Chloride anion exchanger, also known as down-regulated in adenoma, is a protein that in humans is encoded by the SLC26A3 gene.

Solute carrier family 26 member 6 is a protein that in humans is encoded by the SLC26A6 gene. It is an anion-exchanger expressed in the apical membrane of the kidney proximal tubule, the apical membranes of the duct cells in the pancreas, and the villi of the duodenum.

Voltage-dependent anion-selective channel protein 2 is a protein that in humans is encoded by the VDAC2 gene on chromosome 10. This protein is a voltage-dependent anion channel and shares high structural homology with the other VDAC isoforms. VDACs are generally involved in the regulation of cell metabolism, mitochondrial apoptosis, and spermatogenesis. Additionally, VDAC2 participates in cardiac contractions and pulmonary circulation, which implicate it in cardiopulmonary diseases. VDAC2 also mediates immune response to infectious bursal disease (IBD).

miR-96 microRNA precursor is a small non-coding RNA that regulates gene expression. microRNAs are transcribed as ~80 nucleotide precursors and subsequently processed by the Dicer enzyme to give a ~23 nucleotide products. In this case the mature sequence comes from the 5′ arm of the precursor. The mature products are thought to have regulatory roles through complementarity to mRNA.
The cochlear amplifier is a positive feedback mechanism within the cochlea that provides acute sensitivity in the mammalian auditory system. The main component of the cochlear amplifier is the outer hair cell (OHC) which increases the amplitude and frequency selectivity of sound vibrations using electromechanical feedback.

Jonathan Felix Ashmore is a British physicist and Bernard Katz Professor of Biophysics at University College London.
Cochlea is Latin for “snail, shell or screw” and originates from the Greek word κοχλίας kokhlias. The modern definition, the auditory portion of the inner ear, originated in the late 17th century. Within the mammalian cochlea exists the organ of Corti, which contains hair cells that are responsible for translating the vibrations it receives from surrounding fluid-filled ducts into electrical impulses that are sent to the brain to process sound.
Peter Dallos is the John Evans Professor of Neuroscience Emeritus, Professor Emeritus of Audiology, Biomedical Engineering and Otolaryngology at Northwestern University. His research pertained to the neurobiology, biophysics and molecular biology of the cochlea. This work provided the basis for the present understanding of the role of outer hair cells in hearing, that of providing amplification in the cochlea. After his retirement in 2012, he became a professional sculptor.
Hensen's cells are a layer of tall cells arranged in the organ of Corti in the cochlea, which are part of the supporting cells lie on the outer hair cells (OHC). Their appearance are upper part wide with lower part narrow, column like cells. One significant morphologic feature of Hensen's cells is the lipid droplets, which are most noticeable at the third and forth turns of the cochlear, the lipid droplets are thought to have association with the auditory process because they are parallel to the innervation. One significant structure found among the Hensen's cells and the hair cells are the gap junctions, they are made of connexins which serve important function in distribution and connection between cells, the gaps enable the long distance of electric communication.



