
The inner ear is the innermost part of the vertebrate ear. In vertebrates, the inner ear is mainly responsible for sound detection and balance. In mammals, it consists of the bony labyrinth, a hollow cavity in the temporal bone of the skull with a system of passages comprising two main functional parts:

The vestibulocochlear nerve or auditory vestibular nerve, also known as the eighth cranial nerve, cranial nerve VIII, or simply CN VIII, is a cranial nerve that transmits sound and equilibrium (balance) information from the inner ear to the brain. Through olivocochlear fibers, it also transmits motor and modulatory information from the superior olivary complex in the brainstem to the cochlea.

The basilar membrane is a stiff structural element within the cochlea of the inner ear which separates two liquid-filled tubes that run along the coil of the cochlea, the scala media and the scala tympani. The basilar membrane moves up and down in response to incoming sound waves, which are converted to traveling waves on the basilar membrane.

The organ of Corti, or spiral organ, is the receptor organ for hearing and is located in the mammalian cochlea. This highly varied strip of epithelial cells allows for transduction of auditory signals into nerve impulses' action potential. Transduction occurs through vibrations of structures in the inner ear causing displacement of cochlear fluid and movement of hair cells at the organ of Corti to produce electrochemical signals.

The auditory system is the sensory system for the sense of hearing. It includes both the sensory organs and the auditory parts of the sensory system.

An ear is the organ that enables hearing and body balance using the vestibular system. In mammals, the ear is usually described as having three parts: the outer ear, the middle ear and the inner ear. The outer ear consists of the pinna and the ear canal. Since the outer ear is the only visible portion of the ear in most animals, the word "ear" often refers to the external part alone. The middle ear includes the tympanic cavity and the three ossicles. The inner ear sits in the bony labyrinth, and contains structures which are key to several senses: the semicircular canals, which enable balance and eye tracking when moving; the utricle and saccule, which enable balance when stationary; and the cochlea, which enables hearing. The ear canal is cleaned via earwax, which naturally migrates to the auricle. The ears of vertebrates are placed somewhat symmetrically on either side of the head, an arrangement that aids sound localization.
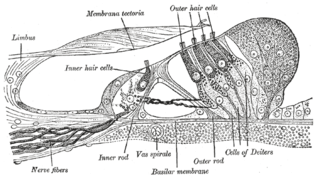
Hair cells are the sensory receptors of both the auditory system and the vestibular system in the ears of all vertebrates, and in the lateral line organ of fishes. Through mechanotransduction, hair cells detect movement in their environment.
Endolymph is the fluid contained in the membranous labyrinth of the inner ear. The major cation in endolymph is potassium, with the values of sodium and potassium concentration in the endolymph being 0.91 mM and 154 mM, respectively. It is also called Scarpa's fluid, after Antonio Scarpa.
In audiology and psychoacoustics the concept of critical bands, introduced by Harvey Fletcher in 1933 and refined in 1940, describes the frequency bandwidth of the "auditory filter" created by the cochlea, the sense organ of hearing within the inner ear. Roughly, the critical band is the band of audio frequencies within which a second tone will interfere with the perception of the first tone by auditory masking.

In the inner ear, stereocilia are the mechanosensing organelles of hair cells, which respond to fluid motion in numerous types of animals for various functions, including hearing and balance. They are about 10–50 micrometers in length and share some similar features of microvilli. The hair cells turn the fluid pressure and other mechanical stimuli into electric stimuli via the many microvilli that make up stereocilia rods. Stereocilia exist in the auditory and vestibular systems.
Perilymph is an extracellular fluid located within the inner ear. It is found within the scala tympani and scala vestibuli of the cochlea. The ionic composition of perilymph is comparable to that of plasma and cerebrospinal fluid. The major cation in perilymph is sodium, with the values of sodium and potassium concentration in the perilymph being 138 mM and 6.9 mM, respectively. It is also named Cotunnius' liquid and liquor cotunnii for Domenico Cotugno.
The tympanic duct or scala tympani is one of the perilymph-filled cavities in the inner ear of humans. It is separated from the cochlear duct by the basilar membrane, and it extends from the round window to the helicotrema, where it continues as vestibular duct.
The cochlear duct is an endolymph filled cavity inside the cochlea, located between the tympanic duct and the vestibular duct, separated by the basilar membrane and the vestibular membrane respectively. The cochlear duct houses the organ of Corti.
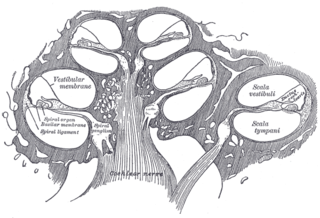
The vestibular membrane, vestibular wall or Reissner's membrane is a membrane inside the cochlea of the inner ear. It separates the cochlear duct from the vestibular duct. It helps to transmit vibrations from fluid in the vestibular duct to the cochlear duct. Together with the basilar membrane, it creates a compartment in the cochlea filled with endolymph, which is important for the function of the spiral organ of Corti. It allows nutrients to travel from the perilymph to the endolymph of the membranous labyrinth. It may be damaged in Ménière's disease. It is named after the German anatomist Ernst Reissner.
An analog ear or analog cochlea is a model of the ear or of the cochlea based on an electrical, electronic or mechanical analog. An analog ear is commonly described as an interconnection of electrical elements such as resistors, capacitors, and inductors; sometimes transformers and active amplifiers are included.
Hearing, or auditory perception, is the ability to perceive sounds through an organ, such as an ear, by detecting vibrations as periodic changes in the pressure of a surrounding medium. The academic field concerned with hearing is auditory science.
The cochlear amplifier is a positive feedback mechanism within the cochlea that provides acute sensitivity in the mammalian auditory system. The main component of the cochlear amplifier is the outer hair cell (OHC) which increases the amplitude and frequency selectivity of sound vibrations using electromechanical feedback.
Electrocochleography is a technique of recording electrical potentials generated in the inner ear and auditory nerve in response to sound stimulation, using an electrode placed in the ear canal or tympanic membrane. The test is performed by an otologist or audiologist with specialized training, and is used for detection of elevated inner ear pressure or for the testing and monitoring of inner ear and auditory nerve function during surgery.
The reticular membrane is a thin, stiff lamina that extends from the outer hair cells to the Hensen's cells. The RM is composed of "minute-fiddle-shaped cuticular structures" called the phalangeal extensions of the outer hair cells, interspaced with extensions coming from the outer phalangeal cells. The RM separates endolymph in the cochlear duct from underlying corticolymph and perilymph of the scala tympani.
Cochlea is Latin for “snail, shell or screw” and originates from the Greek word κοχλίας kokhlias. The modern definition, the auditory portion of the inner ear, originated in the late 17th century. Within the mammalian cochlea exists the organ of Corti, which contains hair cells that are responsible for translating the vibrations it receives from surrounding fluid-filled ducts into electrical impulses that are sent to the brain to process sound.








