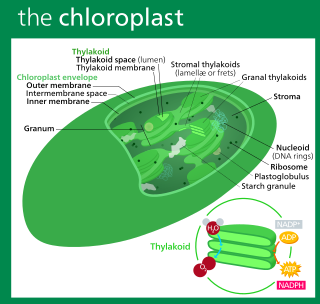
A chloroplast is a type of organelle known as a plastid that conducts photosynthesis mostly in plant and algal cells. Chloroplasts have a high concentration of chlorophyll pigments which capture the energy from sunlight and convert it to chemical energy and release oxygen. The chemical energy created is then used to make sugar and other organic molecules from carbon dioxide in a process called the Calvin cycle. Chloroplasts carry out a number of other functions, including fatty acid synthesis, amino acid synthesis, and the immune response in plants. The number of chloroplasts per cell varies from one, in some unicellular algae, up to 100 in plants like Arabidopsis and wheat.

An endosymbiont or endobiont is an organism that lives within the body or cells of another organism. Typically the two organisms are in a mutualistic relationship. Examples are nitrogen-fixing bacteria, which live in the root nodules of legumes, single-cell algae inside reef-building corals, and bacterial endosymbionts that provide essential nutrients to insects.
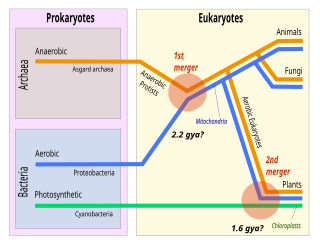
Symbiogenesis is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales bacteria, while chloroplasts are thought to be related to cyanobacteria.
A plastid is a membrane-bound organelle found in the cells of plants, algae, and some other eukaryotic organisms. Plastids are considered to be intracellular endosymbiotic cyanobacteria.
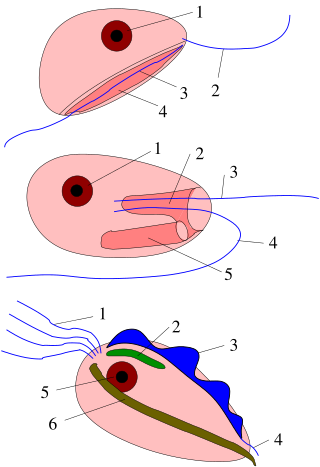
Excavata is an extensive and diverse but paraphyletic group of unicellular Eukaryota. The group was first suggested by Simpson and Patterson in 1999 and the name latinized and assigned a rank by Thomas Cavalier-Smith in 2002. It contains a variety of free-living and symbiotic protists, and includes some important parasites of humans such as Giardia and Trichomonas. Excavates were formerly considered to be included in the now obsolete Protista kingdom. They were distinguished from other lineages based on electron-microscopic information about how the cells are arranged. They are considered to be a basal flagellate lineage.

The glaucophytes, also known as glaucocystophytes or glaucocystids, are a small group of unicellular algae found in freshwater and moist terrestrial environments, less common today than they were during the Proterozoic. The stated number of species in the group varies from about 14 to 26. Together with the red algae (Rhodophyta) and the green algae plus land plants, they form the Archaeplastida.

Chromista is a proposed but polyphyletic biological kingdom, refined from the Chromalveolata, consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.

Nucleomorphs are small, vestigial eukaryotic nuclei found between the inner and outer pairs of membranes in certain plastids. They are thought to be vestiges of primitive red and green algal nuclei that were engulfed by a larger eukaryote. Because the nucleomorph lies between two sets of membranes, nucleomorphs support the endosymbiotic theory and are evidence that the plastids containing them are complex plastids. Having two sets of membranes indicate that the plastid, a prokaryote, was engulfed by a eukaryote, an alga, which was then engulfed by another eukaryote, the host cell, making the plastid an example of secondary endosymbiosis.
In biology, Archezoa is a term that has been introduced by several authors to refer to a group of organisms. Authors include Josef Anton Maximilian Perty, Ernst Haeckel and in the 20th century by Thomas Cavalier-Smith in his classification system. Each author used the name to refer to different arrays of organisms. This reuse by later authors of the same taxon name for different groups of organisms is widely criticized in taxonomy because the inclusion of the name in a sentence does not make sense unless the particular usage is specified. Nonetheless, all uses of 'Archezoa' are now obsolete.

The Archaeplastida are a major group of eukaryotes, comprising the photoautotrophic red algae (Rhodophyta), green algae, land plants, and the minor group glaucophytes. It also includes the non-photosynthetic lineage Rhodelphidia, a predatorial (eukaryotrophic) flagellate that is sister to the Rhodophyta, and probably the microscopic picozoans. The Archaeplastida have chloroplasts that are surrounded by two membranes, suggesting that they were acquired directly through a single endosymbiosis event by phagocytosis of a cyanobacterium. All other groups which have chloroplasts, besides the amoeboid genus Paulinella, have chloroplasts surrounded by three or four membranes, suggesting they were acquired secondarily from red or green algae. Unlike red and green algae, glaucophytes have never been involved in secondary endosymbiosis events.
An apicoplast is a derived non-photosynthetic plastid found in most Apicomplexa, including Toxoplasma gondii, and Plasmodium falciparum and other Plasmodium spp., but not in others such as Cryptosporidium. It originated from algae through secondary endosymbiosis; there is debate as to whether this was a green or red alga. The apicoplast is surrounded by four membranes within the outermost part of the endomembrane system. The apicoplast hosts important metabolic pathways like fatty acid synthesis, isoprenoid precursor synthesis and parts of the heme biosynthetic pathway.
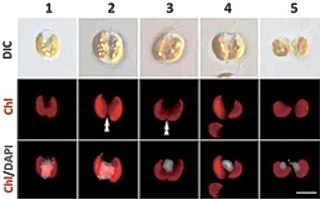
Guillardia is a genus of marine biflagellate cryptomonad algae with a plastid obtained through secondary endosymbiosis of a red alga.

Chloroplast DNA (cpDNA), also known as plastid DNA (ptDNA) is the DNA located in chloroplasts, which are photosynthetic organelles located within the cells of some eukaryotic organisms. Chloroplasts, like other types of plastid, contain a genome separate from that in the cell nucleus. The existence of chloroplast DNA was identified biochemically in 1959, and confirmed by electron microscopy in 1962. The discoveries that the chloroplast contains ribosomes and performs protein synthesis revealed that the chloroplast is genetically semi-autonomous. The first complete chloroplast genome sequences were published in 1986, Nicotiana tabacum (tobacco) by Sugiura and colleagues and Marchantia polymorpha (liverwort) by Ozeki et al. Since then, tens of thousands of chloroplast genomes from various species have been sequenced.

Cyanidiophyceae is a class of unicellular red algae within subdivision Cyanidiophytina, and contain a single plastid, one to three mitochondria, a nucleus, a vacuole, and floridean starch. Pyrenoids are absent. Most are extremophiles inhabiting acid hot springs. They originated in extreme environments with high temperatures and low pH, which allowed them to occupy ecological niches without any competition. While still found in extreme environments, they have also adapted to live along streams, in fissures in rock walls and in soil, but usually prefer relatively high temperatures. They have never been found in basic freshwater or seawater habitats. The main photosynthetic pigment is C-phycocyanin. Reproduction is asexual by binary fission or formation of endospores. The group, consisting of a single order (Cyanidiales), split off from the other red algae more than a billion years ago. Three families, four genera, and nine species are known, but the total number of species is probably higher. They are primarily photoautotrophic, but heterotrophic and mixotrophic growth also occurs. After the first massive gene loss in the common ancestor of all red algae, where ca. 25% of the genes were lost, a second gene loss occurred in the ancestor of Cyanidiophyceae, where additional 18% of the genes were lost. Since then, some gene gains and minor gene losses have taken place independently in the Cyanidiaceae and Galdieriaceae, leading to genetic diversification between the two groups, with Galdieriaceae occupying more diverse and varied niches in extreme environments than Cyanidiaceae.

Picozoa, Picobiliphyta, Picobiliphytes, or Biliphytes are protists of a phylum of marine unicellular heterotrophic eukaryotes with a size of less than about 3 micrometers. They were formerly treated as eukaryotic algae and the smallest member of photosynthetic picoplankton before it was discovered they do not perform photosynthesis. The first species identified therein is Picomonas judraskeda. They probably belong in the Archaeplastida as sister of the Rhodophyta.

Mesodinium rubrum is a species of ciliates. It constitutes a plankton community and is found throughout the year, most abundantly in spring and fall, in coastal areas. Although discovered in 1908, its scientific importance came into light in the late 1960s when it attracted scientists by the recurrent red colouration it caused by forming massive blooms, that cause red tides in the oceans.

Floridean starch is a type of a storage glucan found in glaucophytes and in red algae, in which it is usually the primary sink for fixed carbon from photosynthesis. It is found in grains or granules in the cell's cytoplasm and is composed of an α-linked glucose polymer with a degree of branching intermediate between amylopectin and glycogen, though more similar to the former. The polymers that make up floridean starch are sometimes referred to as "semi-amylopectin".

Gloeomargarita lithophora, a cyanobacterium, is the proposed closest present day relative of all chloroplasts. The ancient relative of Gloeomargarita's was engulfed by a eukaryotic host in a singule endosymbiotic event around 1900-1400 million years ago. The origin of plastids by endosymbiosis signifies the beginning of photosynthesis in eukaryotes, and as such their evolutionary relationship to Gloeomargarita lithophora, as the sister group, is of high importance to the evolutionary history of endosymbiotic organelles and photosynthesis.
A plastid is a membrane-bound organelle found in plants, algae and other eukaryotic organisms that contribute to the production of pigment molecules. Most plastids are photosynthetic, thus leading to color production and energy storage or production. There are many types of plastids in plants alone, but all plastids can be separated based on the number of times they have undergone endosymbiotic events. Currently there are three types of plastids; primary, secondary and tertiary. Endosymbiosis is reputed to have led to the evolution of eukaryotic organisms today, although the timeline is highly debated.

Photoautotrophs are organisms that can utilize light energy from sunlight and elements from inorganic compounds to produce organic materials needed to sustain their own metabolism. Such biological activities are known as photosynthesis, and examples of such organisms include plants, algae and cyanobacteria.
