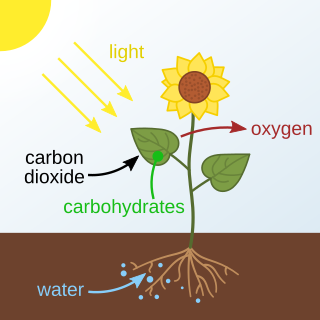
Photosynthesis is a system of biological processes by which photosynthetic organisms, such as most plants, algae, and cyanobacteria, convert light energy, typically from sunlight, into the chemical energy necessary to fuel their metabolism. Photosynthesis usually refers to oxygenic photosynthesis, a process that produces oxygen. Photosynthetic organisms store the chemical energy so produced within intracellular organic compounds like sugars, glycogen, cellulose and starches. To use this stored chemical energy, an organism's cells metabolize the organic compounds through cellular respiration. Photosynthesis plays a critical role in producing and maintaining the oxygen content of the Earth's atmosphere, and it supplies most of the biological energy necessary for complex life on Earth.

The green sulfur bacteria are a phylum, Chlorobiota, of obligately anaerobic photoautotrophic bacteria that metabolize sulfur.

Chloroflexus aurantiacus is a photosynthetic bacterium isolated from hot springs, belonging to the green non-sulfur bacteria. This organism is thermophilic and can grow at temperatures from 35 to 70 °C. Chloroflexus aurantiacus can survive in the dark if oxygen is available. When grown in the dark, Chloroflexus aurantiacus has a dark orange color. When grown in sunlight it is dark green. The individual bacteria tend to form filamentous colonies enclosed in sheaths, which are known as trichomes.
The purple sulfur bacteria (PSB) are part of a group of Pseudomonadota capable of photosynthesis, collectively referred to as purple bacteria. They are anaerobic or microaerophilic, and are often found in stratified water environments including hot springs, stagnant water bodies, as well as microbial mats in intertidal zones. Unlike plants, algae, and cyanobacteria, purple sulfur bacteria do not use water as their reducing agent, and therefore do not produce oxygen. Instead, they can use sulfur in the form of sulfide, or thiosulfate (as well, some species can use H2, Fe2+, or NO2−) as the electron donor in their photosynthetic pathways. The sulfur is oxidized to produce granules of elemental sulfur. This, in turn, may be oxidized to form sulfuric acid.
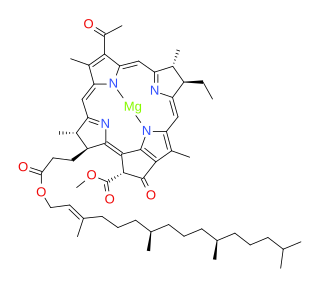
Bacteriochlorophylls (BChl) are photosynthetic pigments that occur in various phototrophic bacteria. They were discovered by C. B. van Niel in 1932. They are related to chlorophylls, which are the primary pigments in plants, algae, and cyanobacteria. Organisms that contain bacteriochlorophyll conduct photosynthesis to sustain their energy requirements, but the process is anoxygenic and does not produce oxygen as a byproduct. They use wavelengths of light not absorbed by plants or cyanobacteria. Replacement of Mg2+ with protons gives bacteriophaeophytin (BPh), the phaeophytin form.
Heliobacteria are a unique subset of prokaryotic bacteria that process light for energy. Distinguishable from other phototrophic bacteria, they utilize a unique photosynthetic pigment, bacteriochlorophyll g and are the only known Gram-positive phototroph. They are a key player in symbiotic nitrogen fixation alongside plants, and use a type I reaction center like green-sulfur bacteria.

The Chromatiaceae are one of the two families of purple sulfur bacteria, together with the Ectothiorhodospiraceae. They belong to the order Chromatiales of the class Gammaproteobacteria, which is composed by unicellular Gram-negative organisms. Most of the species are photolithoautotrophs and conduct an anoxygenic photosynthesis, but there are also representatives capable of growing under dark and/or microaerobic conditions as either chemolithoautotrophs or chemoorganoheterotrophs.
Photoheterotrophs are heterotrophic phototrophs—that is, they are organisms that use light for energy, but cannot use carbon dioxide as their sole carbon source. Consequently, they use organic compounds from the environment to satisfy their carbon requirements; these compounds include carbohydrates, fatty acids, and alcohols. Examples of photoheterotrophic organisms include purple non-sulfur bacteria, green non-sulfur bacteria, and heliobacteria. These microorganisms are ubiquitous in aquatic habitats, occupy unique niche-spaces, and contribute to global biogeochemical cycling. Recent research has also indicated that the oriental hornet and some aphids may be able to use light to supplement their energy supply.

A chlorosome is a photosynthetic antenna complex found in green sulfur bacteria (GSB) and many green non-sulfur bacteria (GNsB), together known as green bacteria. They differ from other antenna complexes by their large size and lack of protein matrix supporting the photosynthetic pigments. Green sulfur bacteria are a group of organisms that generally live in extremely low-light environments, such as at depths of 100 metres in the Black Sea. The ability to capture light energy and rapidly deliver it to where it needs to go is essential to these bacteria, some of which see only a few photons of light per chlorophyll per day. To achieve this, the bacteria contain chlorosome structures, which contain up to 250,000 chlorophyll molecules. Chlorosomes are ellipsoidal bodies, in GSB their length varies from 100 to 200 nm, width of 50-100 nm and height of 15 – 30 nm, in GNsB the chlorosomes are somewhat smaller.
γ-Carotene (gamma-carotene) is a carotenoid, and is a biosynthetic intermediate for cyclized carotenoid synthesis in plants. It is formed from cyclization of lycopene by lycopene cyclase epsilon. Along with several other carotenoids, γ-carotene is a vitamer of vitamin A in herbivores and omnivores. Carotenoids with a cyclized, beta-ionone ring can be converted to vitamin A, also known as retinol, by the enzyme beta-carotene 15,15'-dioxygenase; however, the bioconversion of γ-carotene to retinol has not been well-characterized. γ-Carotene has tentatively been identified as a biomarker for green and purple sulfur bacteria in a sample from the 1.640 ± 0.003-Gyr-old Barney Creek Formation in Northern Australia which comprises marine sediments. Tentative discovery of γ-carotene in marine sediments implies a past euxinic environment, where water columns were anoxic and sulfidic. This is significant for reconstructing past oceanic conditions, but so far γ-carotene has only been potentially identified in the one measured sample.
Sulfur is metabolized by all organisms, from bacteria and archaea to plants and animals. Sulfur can have an oxidation state from -2 to +6 and is reduced or oxidized by a diverse range of organisms. The element is present in proteins, sulfate esters of polysaccharides, steroids, phenols, and sulfur-containing coenzymes.
Rhodopseudomonas palustris is a rod-shaped, Gram-negative purple nonsulfur bacterium, notable for its ability to switch between four different modes of metabolism.

Anoxygenic photosynthesis is a special form of photosynthesis used by some bacteria and archaea, which differs from the better known oxygenic photosynthesis in plants in the reductant used and the byproduct generated.
Rhodovulum sulfidophilum is a gram-negative purple nonsulfur bacteria. The cells are rod-shaped, and range in size from 0.6 to 0.9 μm wide and 0.9 to 2.0 μm long, and have a polar flagella. These cells reproduce asexually by binary fission. This bacterium can grow anaerobically when light is present, or aerobically (chemoheterotrophic) under dark conditions. It contains the photosynthetic pigments bacteriochlorophyll a and of carotenoids.
Chlorobaculum tepidum, previously known as Chlorobium tepidum, is an anaerobic, thermophilic green sulfur bacteria first isolated from New Zealand. Its cells are gram-negative and non-motile rods of variable length. They contain chlorosomes and bacteriochlorophyll a and c.
Chlorobium chlorochromatii, originally known as Chlorobium aggregatum, is a symbiotic green sulfur bacteria that performs anoxygenic photosynthesis and functions as an obligate photoautotroph using reduced sulfur species as electron donors. Chlorobium chlorochromatii can be found in stratified freshwater lakes.
Okenane, the diagenetic end product of okenone, is a biomarker for Chromatiaceae, the purple sulfur bacteria. These anoxygenic phototrophs use light for energy and sulfide as their electron donor and sulfur source. Discovery of okenane in marine sediments implies a past euxinic environment, where water columns were anoxic and sulfidic. This is potentially tremendously important for reconstructing past oceanic conditions, but so far okenane has only been identified in one Paleoproterozoic rock sample from Northern Australia.

Microbial oxidation of sulfur is the oxidation of sulfur by microorganisms to build their structural components. The oxidation of inorganic compounds is the strategy primarily used by chemolithotrophic microorganisms to obtain energy to survive, grow and reproduce. Some inorganic forms of reduced sulfur, mainly sulfide (H2S/HS−) and elemental sulfur (S0), can be oxidized by chemolithotrophic sulfur-oxidizing prokaryotes, usually coupled to the reduction of oxygen (O2) or nitrate (NO3−). Anaerobic sulfur oxidizers include photolithoautotrophs that obtain their energy from sunlight, hydrogen from sulfide, and carbon from carbon dioxide (CO2).
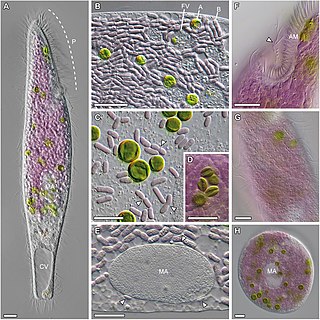
Pseudoblepharisma is a genus of heterotrich ciliates inhabiting oxygen depleted freshwater habitats. Most sources report that it contains one species, Pseudoblepharisma tenue, but at least four have been seen in literature.

Prosthecochloris aestuarii is a green sulfur bacterium in the genus Prosthecochloris. This organism was originally isolated from brackish lagoons located in Sasyk-Sivash and Sivash. They are characterized by the presence of "prosthecae" on their cell surface; the inner part of these appendages house the photosynthetic machinery within chlorosomes, which are characteristic structures of green sulfur bacteria. Additionally, like other green sulfur bacteria, they are Gram-negative, non-motile, and non-spore forming. Of the four major groups of green sulfur bacteria, P. aestuarii serves as the type species for Group 4.

