| Mammalian kidney | |
|---|---|
 | |
| Details | |
| Precursor | Ureteric bud, metanephrogenic blastema |
| System | Urinary system and endocrine system |
| Artery | Renal artery |
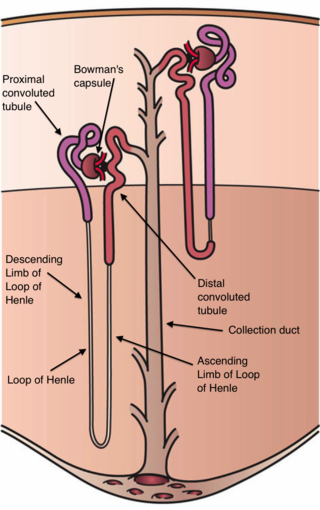
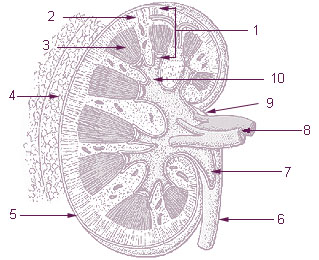
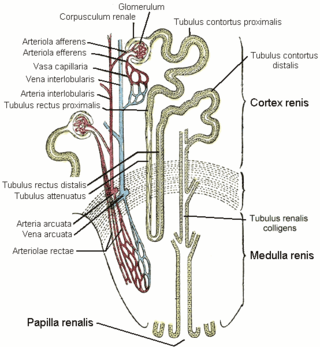
| Vein | Renal vein |
| Nerve | Renal plexus |
| Lymph | Collecting lymphatic vessels |
| Anatomical terminology | |
The mammalian kidneys are a pair of excretory organs of the urinary system of mammals, [2] being functioning kidneys in postnatal-to-adult individuals [3] (i. e. metanephric kidneys). [2] The kidneys in mammals are usually bean-shaped [4] or externally lobulated. [5] They are located behind the peritoneum (retroperitoneally) [6] on the back (dorsal) wall of the body. [7] The typical mammalian kidney consists of a renal capsule, a peripheral cortex, an internal medulla, one or more renal calyces, and a renal pelvis. [7] Although the calyces or renal pelvis may be absent in some species. [7] The medulla is made up of one or more renal pyramids, [8] forming papillae with their innermost parts. [9] Generally, urine produced by the cortex and medulla drains from the papillae into the calyces, and then into the renal pelvis, from which urine exits the kidney through the ureter. [7] [10] Nitrogen-containing waste products are excreted by the kidneys in mammals mainly in the form of urea. [11]
Contents
- Structure
- Gross anatomy
- Parenchyma
- Variations
- Microanatomy
- Blood supply
- Lymphatic drainage
- Nerve supply
- Functions
- Excretory function
- Osmoregulation
- Endocrine function
- Blood pressure regulation
- Acid-base balance
- Glucose homeostasis
- Evolution
- Mammalian metanephric kidney
- Adaptations to aridity
- Adaptations to body mass
- Reniculate kidneys
- Development
- Stages of kidney development
- Metanephros development
- Postnatal maturation
- Injury and diseases
- Congenital and inherited anomalies
- Non-infectious diseases
- Infectious diseases
- Ageing
- Repair and regeneration
- Compensatory capabilities
- Nephron regeneration
- Healing after injury
- See also
- References
- Bibliography
- External links
The structure of the kidney differs between species. [12] The kidneys can be unilobar (a single lobe represented by a single renal pyramid) or multilobar, [13] [14] unipapillary (a single or a common papilla), with several papillae or multipapillary, [14] [15] may be smooth-surfaced or lobulated. [1] [13] The multilobar kidneys can also be reniculate, which are found mainly in marine mammals. [16] The unipapillary kidney with a single renal pyramid is the simplest type of kidney in mammals, from which the more structurally complex kidneys are believed to have evolved. [17] [6] [18] Differences in kidney structure are the result of adaptations during evolution to variations in body mass and habitats (in particular, aridity) between species. [19] [20] [12]
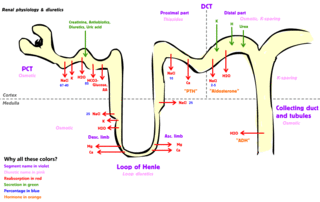
The cortex and medulla of the kidney contain nephrons, [21] each of which consists of a glomerulus and a complex tubular system. [22] The cortex contains glomeruli and is responsible for filtering the blood. [7] The medulla is responsible for urine concentration [23] and contains tubules with short and long loops of Henle. [24] The loops of Henle are essential for urine concentration. [25] Amongst the vertebrates, only mammals and birds have kidneys that can produce urine more concentrated (hypertonic) than the blood plasma, [7] but only in mammals do all nephrons have the loop of Henle. [26]
The kidneys of mammals are vital organs [27] that maintain water, electrolyte and acid-base balance in the body, excrete nitrogenous waste products, regulate blood pressure, and participate in bone formation [28] [29] [30] and regulation of glucose levels. [31] The processes of blood plasma filtration, tubular reabsorption and tubular secretion occur in the kidneys, and urine formation is a result of these processes. [8] The kidneys produce renin [32] and erythropoietin [33] hormones, and are involved in the conversion of vitamin D to its active form. [34] Mammals are the only class of vertebrates in which only the kidneys are responsible for maintaining the homeostasis of the extracellular fluid in the body. [35] The function of the kidneys is regulated by the autonomic nervous system and hormones. [36]
The potential for regeneration in mature kidneys is limited [37] [38] because new nephrons cannot be formed. [39] But in cases of limited injury, renal function can be restored through compensatory mechanisms. [40] The kidneys can have noninfectious and infectious diseases; in rare cases, congenital and hereditary anomalies occur in the kidneys of mammals. [41] Pyelonephritis is usually caused by bacterial infections. [42] [43] Some diseases may be species specific, [44] and parasitic kidney diseases are common in some species. [45] [46] The structural characteristics of the mammalian kidneys make them vulnerable to ischemic and toxic injuries. [47] Permanent damage can lead to chronic kidney disease. [48] [49] Ageing of the kidneys also causes changes in them, and the number of functioning nephrons decreases with age. [50]