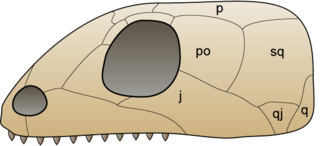
An anapsid is an amniote whose skull lacks one or more skull openings near the temples. Traditionally, the Anapsida are considered the most primitive subclass of amniotes, the ancestral stock from which Synapsida and Diapsida evolved, making anapsids paraphyletic. It is, however, doubtful that all anapsids lack temporal fenestra as a primitive trait, and that all the groups traditionally seen as anapsids truly lacked fenestra.

Amniotes are tetrapod vertebrate animals belonging to the clade Amniota, a large group that comprises the vast majority of living terrestrial and semiaquatic vertebrates. Amniotes evolved from amphibian ancestors during the Carboniferous period and further diverged into two groups, namely the sauropsids and synapsids, an event that marks the appearance of Amniota, according to the definition established under the PhyloCode. This basal divergence within Amniota has been dated by molecular studies at 310–329 Ma or 312–330 Ma, but the presence of Hylonomus at Joggins implies a minimal age of about 317 Ma. A fossilized birth-death process study of early amniotes suggested an age of 322–340 Ma. Amniotes are distinguished from the other living tetrapod clade — the non-amniote lissamphibians — by the development of three extraembryonic membranes, thicker and keratinized skin, and costal respiration. Additional unique features are the presence of adrenocortical and chromaffin tissues as a discrete pair of glands near their kidneys, which are more complex, the presence of an astragalus for better extremity range of motion, the diminished role of skin breathing, and the complete loss of metamorphosis, gills, and lateral lines.

The Lepidosauria is a subclass or superorder of reptiles, containing the orders Squamata and Rhynchocephalia. Squamata includes lizards and snakes. Squamata contains over 9,000 species, making it by far the most species-rich and diverse order of non-avian reptiles in the present day. Rhynchocephalia was a formerly widespread and diverse group of reptiles in the Mesozoic Era. However, it is represented by only one living species: the tuatara, a superficially lizard-like reptile native to New Zealand.
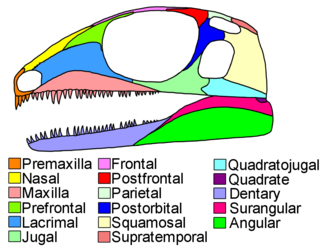
Diapsids are a clade of sauropsids, distinguished from more primitive eureptiles by the presence of two holes, known as temporal fenestrae, in each side of their skulls. The earliest traditionally identified diapsids, the araeoscelidians, appeared about three hundred million years ago during the late Carboniferous period. All diapsids other than the most primitive ones in the clade Araeoscelidia are often placed into the clade Neodiapsida. The diapsids are extremely diverse, and include birds and all modern reptile groups, including turtles, which were historically thought to lie outside the group. All modern reptiles and birds are placed within the neodiapsid subclade Sauria. Although some diapsids have lost either one hole (lizards), or both holes, or have a heavily restructured skull, they are still classified as diapsids based on their ancestry. At least 17,084 species of diapsid animals are extant: 9,159 birds, and 7,925 snakes, lizards, tuatara, turtles, and crocodiles.

Sauropsida is a clade of amniotes, broadly equivalent to the class Reptilia, though typically used in a broader sense to also include extinct stem-group relatives of modern reptiles and birds. The most popular definition states that Sauropsida is the sibling taxon to Synapsida, the other clade of amniotes which includes mammals as its only modern representatives. Although early synapsids have historically been referred to as "mammal-like reptiles", all synapsids are more closely related to mammals than to any modern reptile. Sauropsids, on the other hand, include all amniotes more closely related to modern reptiles than to mammals. This includes Aves (birds), which are recognized as a subgroup of archosaurian reptiles despite originally being named as a separate class in Linnaean taxonomy.

Sauropterygia is an extinct taxon of diverse, aquatic reptiles that developed from terrestrial ancestors soon after the end-Permian extinction and flourished during the Triassic before all except for the Plesiosauria became extinct at the end of that period. The plesiosaurs would continue to diversify until the end of the Mesozoic. Sauropterygians are united by a radical adaptation of their pectoral girdle, adapted to support powerful flipper strokes. Some later sauropterygians, such as the pliosaurs, developed a similar mechanism in their pelvis. It is possible that sauropterygians are a distant relatives of turtles, uniting them under the group Pantestudines, although this is still debatable as sauropterygians might be archosauromorphs or completely unrelated to both.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.

Eureptilia is one of the two major subgroups of the clade Sauropsida, the other one being Parareptilia. Eureptilia includes Diapsida, as well as a number of primitive Permo-Carboniferous forms previously classified under Anapsida, in the old order "Cotylosauria".
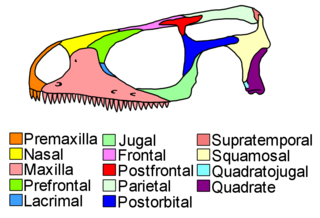
Lepidosauromorpha is a group of reptiles comprising all diapsids closer to lizards than to archosaurs. The only living sub-group is the Lepidosauria, which contains two subdivisions, Squamata, which contains lizards and snakes, and Rhynchocephalia, the only extant species of which is the tuatara.
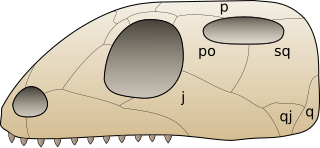
Euryapsida is a polyphyletic group of sauropsids that are distinguished by a single temporal fenestra, an opening behind the orbit, under which the post-orbital and squamosal bones articulate. They are different from Synapsida, which also have a single opening behind the orbit, by the placement of the fenestra. In synapsids, this opening is below the articulation of the post-orbital and squamosal bones. It is now commonly believed that euryapsids are in fact diapsids that lost the lower temporal fenestra. Euryapsids are usually considered entirely extinct, although turtles might be part of the sauropterygian clade while other authors disagree. Euryapsida may also be a synonym of Sauropterygia sensu lato.

Avicephala is a potentially polyphyletic grouping of extinct diapsid reptiles that lived during the Late Permian and Triassic periods characterised by superficially bird-like skulls and arboreal lifestyles. As a clade, Avicephala is defined as including the gliding weigeltisaurids and the arboreal drepanosaurs to the exclusion of other major diapsid groups. This relationship is not recovered in the majority of phylogenetic analyses of early diapsids and so Avicephala is typically regarded as an artificial or unnatural grouping. However, the clade was recovered again in 2021 following a redescription of Weigeltisaurus, raising the possibility that the clade may be valid after all, although subsequent analyses have not supported this result.

Parareptilia ("near-reptiles") is an extinct subclass or clade of basal sauropsids/reptiles, typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.

Younginiformes is a group of diapsid reptiles known from the Permian-Triassic of Africa and Madagascar. It has been used as a replacement for "Eosuchia". Younginiformes were historically suggested to be lepidosauromorphs, but are currently thought to be basal non-saurian neodiapsids. The group is sometimes divided into two families, Tangasauridae and Younginidae. The monophyly of the group is disputed. A 2009 study found them to be an unresolved polytomy at the base of Neodiapsida, while a 2011 study recovered the group as paraphyletic. A 2022 study recovered the Younginiformes as a monophyletic group of basal neodiapsid reptiles, also including Claudiosaurus and Saurosternon as part of the group. Some younginiforms like Hovasaurus and Acerosodontosaurus are thought to have had an amphibious lifestyle, while others like Kenyasaurus, Thadeosaurus and Youngina were probably terrestrial.

Thalattosauria is an extinct order of marine reptiles that lived in the Middle to Late Triassic. Thalattosaurs were diverse in size and shape, and are divided into two superfamilies: Askeptosauroidea and Thalattosauroidea. Askeptosauroids were endemic to the Tethys Ocean, their fossils have been found in Europe and China, and they were likely semiaquatic fish eaters with straight snouts and decent terrestrial abilities. Thalattosauroids were more specialized for aquatic life and most had unusual downturned snouts and crushing dentition. Thalattosauroids lived along the coasts of both Panthalassa and the Tethys Ocean, and were most diverse in China and western North America. The largest species of thalattosaurs grew to over 4 meters (13 feet) in length, including a long, flattened tail utilized in underwater propulsion. Although thalattosaurs bore a superficial resemblance to lizards, their exact relationships are unresolved. They are widely accepted as diapsids, but experts have variously placed them on the reptile family tree among Lepidosauromorpha, Archosauromorpha, ichthyosaurs, and/or other marine reptiles.

Acerosodontosaurus is an extinct genus of neodiapsid reptiles that lived during the Late Permian of Madagascar. The only species of Acerosodontosaurus, A. piveteaui, is known from a natural mold of a single partial skeleton including a crushed skull and part of the body and limbs. The fossil was discovered in deposits of the Lower Sakamena Formation. Based on skeletal characteristics, it has been suggested that Acerosodontosaurus individuals were at least partially aquatic.

Eunotosaurus is an extinct genus of amniote, possibly a close relative of turtles. Eunotosaurus lived in the late Middle Permian and fossils can be found in the Karoo Supergroup of South Africa and Malawi. Eunotosaurus resided in the swamps of what is now southern Africa. Its ribs were wide and flat, forming broad plates similar to a primitive turtle shell, and the vertebrae were nearly identical to those of some turtles. Accordingly, it is often considered as a possible transitional fossil between turtles and their prehistoric ancestors. However, it is possible that these turtle-like features evolved independently of the same features in turtles, since other anatomical studies and phylogenetic analyses suggest that Eunotosaurus may instead have been a parareptile, an early-diverging neodiapsid unrelated to turtles, or a synapsid.

Archelosauria is a clade grouping turtles and archosaurs and their fossil relatives, to the exclusion of lepidosaurs. The majority of phylogenetic analyses based on molecular data have supported a sister-group relationship between turtles and archosaurs. On the other hand, Archelosauria had not been historically supported by most morphological analyses, which have instead found turtles to either be descendants of parareptiles, early-diverging diapsids outside of Sauria, or close relatives of lepidosaurs within the clade Ankylopoda. Some recent morphological analyses have also found support for Archelosauria.

Pantestudines or Pan-Testudines is the proposed group of all reptiles more closely related to turtles than to any other living animal. It includes both modern turtles and all of their extinct relatives. Pantestudines with a complete shell are placed in the clade Testudinata.

Pappochelys is an extinct genus of diapsid reptile possibly related to turtles. The genus contains only one species, Pappochelys rosinae, from the Middle Triassic of Germany, which was named by paleontologists Rainer Schoch and Hans-Dieter Sues in 2015. The discovery of Pappochelys provides strong support for the placement of turtles within Diapsida, a hypothesis that has long been suggested by molecular data, but never previously by the fossil record. It is morphologically intermediate between the definite stem-turtle Odontochelys from the Late Triassic of China and Eunotosaurus, a reptile from the Middle Permian of South Africa.
Ankylopoda was a proposed clade that hypothetically contains turtles and lepidosaurs and their fossil relatives. This clade was historically supported based on microRNA analysis as well as some cladistic analyses. However, it was strongly contradicted by molecular evidence which supports Archelosauria, and other recent cladistic analyses have supported Archelosauria over Ankylopoda.









