An allele, or allelomorph, is a variant of the sequence of nucleotides at a particular location, or locus, on a DNA molecule.
The genotype of an organism is its complete set of genetic material. Genotype can also be used to refer to the alleles or variants an individual carries in a particular gene or genetic location. The number of alleles an individual can have in a specific gene depends on the number of copies of each chromosome found in that species, also referred to as ploidy. In diploid species like humans, two full sets of chromosomes are present, meaning each individual has two alleles for any given gene. If both alleles are the same, the genotype is referred to as homozygous. If the alleles are different, the genotype is referred to as heterozygous.
Heredity, also called inheritance or biological inheritance, is the passing on of traits from parents to their offspring; either through asexual reproduction or sexual reproduction, the offspring cells or organisms acquire the genetic information of their parents. Through heredity, variations between individuals can accumulate and cause species to evolve by natural selection. The study of heredity in biology is genetics.

Mendelian inheritance is a type of biological inheritance following the principles originally proposed by Gregor Mendel in 1865 and 1866, re-discovered in 1900 by Hugo de Vries and Carl Correns, and later popularized by William Bateson. These principles were initially controversial. When Mendel's theories were integrated with the Boveri–Sutton chromosome theory of inheritance by Thomas Hunt Morgan in 1915, they became the core of classical genetics. Ronald Fisher combined these ideas with the theory of natural selection in his 1930 book The Genetical Theory of Natural Selection, putting evolution onto a mathematical footing and forming the basis for population genetics within the modern evolutionary synthesis.
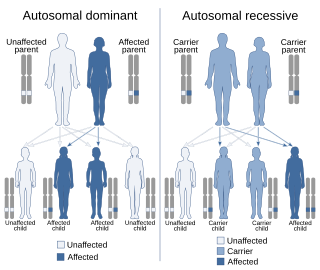
In genetics, dominance is the phenomenon of one variant (allele) of a gene on a chromosome masking or overriding the effect of a different variant of the same gene on the other copy of the chromosome. The first variant is termed dominant and the second is called recessive. This state of having two different variants of the same gene on each chromosome is originally caused by a mutation in one of the genes, either new or inherited. The terms autosomal dominant or autosomal recessive are used to describe gene variants on non-sex chromosomes (autosomes) and their associated traits, while those on sex chromosomes (allosomes) are termed X-linked dominant, X-linked recessive or Y-linked; these have an inheritance and presentation pattern that depends on the sex of both the parent and the child. Since there is only one copy of the Y chromosome, Y-linked traits cannot be dominant or recessive. Additionally, there are other forms of dominance, such as incomplete dominance, in which a gene variant has a partial effect compared to when it is present on both chromosomes, and co-dominance, in which different variants on each chromosome both show their associated traits.
Genetic linkage is the tendency of DNA sequences that are close together on a chromosome to be inherited together during the meiosis phase of sexual reproduction. Two genetic markers that are physically near to each other are unlikely to be separated onto different chromatids during chromosomal crossover, and are therefore said to be more linked than markers that are far apart. In other words, the nearer two genes are on a chromosome, the lower the chance of recombination between them, and the more likely they are to be inherited together. Markers on different chromosomes are perfectly unlinked, although the penetrance of potentially deleterious alleles may be influenced by the presence of other alleles, and these other alleles may be located on other chromosomes than that on which a particular potentially deleterious allele is located.
A heterozygote advantage describes the case in which the heterozygous genotype has a higher relative fitness than either the homozygous dominant or homozygous recessive genotype. Loci exhibiting heterozygote advantage are a small minority of loci. The specific case of heterozygote advantage due to a single locus is known as overdominance. Overdominance is a rare condition in genetics where the phenotype of the heterozygote lies outside of the phenotypical range of both homozygote parents, and heterozygous individuals have a higher fitness than homozygous individuals.

Equine coat color genetics determine a horse's coat color. Many colors are possible, but all variations are produced by changes in only a few genes. Bay is the most common color of horse, followed by black and chestnut. A change at the agouti locus is capable of turning bay to black, while a mutation at the extension locus can turn bay or black to chestnut.
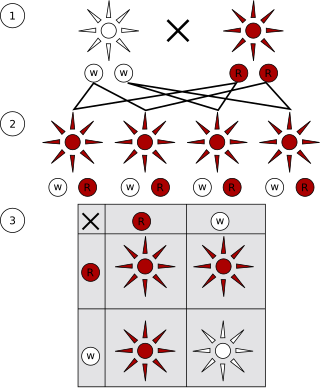
A monohybrid cross is a cross between two organisms with different variations at one genetic locus of interest. The character(s) being studied in a monohybrid cross are governed by two or multiple variations for a single location of a gene. Then carry out such a cross, each parent is chosen to be homozygous or true breeding for a given trait (locus). When a cross satisfies the conditions for a monohybrid cross, it is usually detected by a characteristic distribution of second-generation (F2) offspring that is sometimes called the monohybrid ratio.

Non-Mendelian inheritance is any pattern in which traits do not segregate in accordance with Mendel's laws. These laws describe the inheritance of traits linked to single genes on chromosomes in the nucleus. In Mendelian inheritance, each parent contributes one of two possible alleles for a trait. If the genotypes of both parents in a genetic cross are known, Mendel's laws can be used to determine the distribution of phenotypes expected for the population of offspring. There are several situations in which the proportions of phenotypes observed in the progeny do not match the predicted values.
Human genetics is the study of inheritance as it occurs in human beings. Human genetics encompasses a variety of overlapping fields including: classical genetics, cytogenetics, molecular genetics, biochemical genetics, genomics, population genetics, developmental genetics, clinical genetics, and genetic counseling.
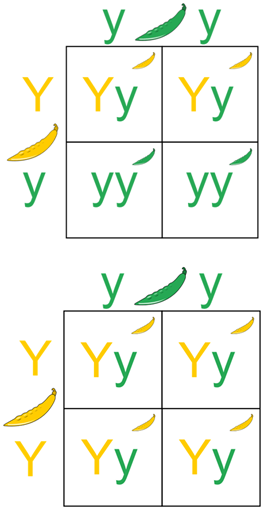
Dihybrid cross is a cross between two individuals with two observed traits that are controlled by two distinct genes. The idea of a dihybrid cross came from Gregor Mendel when he observed pea plants that were either yellow or green and either round or wrinkled. Crossing of two heterozygous individuals will result in predictable ratios for both genotype and phenotype in the offspring. The expected phenotypic ratio of crossing heterozygous parents would be 9:3:3:1. Deviations from these expected ratios may indicate that the two traits are linked or that one or both traits has a non-Mendelian mode of inheritance.
In genetics, a reciprocal cross is a breeding experiment designed to test the role of parental sex on a given inheritance pattern. All parent organisms must be true breeding to properly carry out such an experiment. In one cross, a male expressing the trait of interest will be crossed with a female not expressing the trait. In the other, a female expressing the trait of interest will be crossed with a male not expressing the trait. It is the cross that could be made either way or independent of the sex of the parents. For example, suppose a biologist wished to identify whether a hypothetical allele Z, a variant of some gene A, is on the male or female sex chromosome. They might first cross a Z-trait female with an A-trait male and observe the offspring. Next, they would cross an A-trait female with a Z-trait male and observe the offspring. Via principles of dominant and recessive alleles, they could then make an inference as to which sex chromosome contains the gene Z, if either in fact did.
Under the law of dominance in genetics, an individual expressing a dominant phenotype could contain either two copies of the dominant allele or one copy of each dominant and recessive allele. By performing a test cross, one can determine whether the individual is heterozygous or homozygous dominant.
Out-crossing or out-breeding is the technique of crossing between different breeds. This is the practice of introducing distantly related genetic material into a breeding line, thereby increasing genetic diversity.
Lethal alleles are alleles that cause the death of the organism that carries them. They are usually a result of mutations in genes that are essential for growth or development. Lethal alleles may be recessive, dominant, or conditional depending on the gene or genes involved.

The genetic basis of coat colour in the Labrador Retriever has been found to depend on several distinct genes. The interplay among these genes is used as an example of epistasis.

Particulate inheritance is a pattern of inheritance discovered by Mendelian genetics theorists, such as William Bateson, Ronald Fisher or Gregor Mendel himself, showing that phenotypic traits can be passed from generation to generation through "discrete particles" known as genes, which can keep their ability to be expressed while not always appearing in a descending generation.
Classical genetics is the branch of genetics based solely on visible results of reproductive acts. It is the oldest discipline in the field of genetics, going back to the experiments on Mendelian inheritance by Gregor Mendel who made it possible to identify the basic mechanisms of heredity. Subsequently, these mechanisms have been studied and explained at the molecular level.
This glossary of genetics and evolutionary biology is a list of definitions of terms and concepts used in the study of genetics and evolutionary biology, as well as sub-disciplines and related fields, with an emphasis on classical genetics, quantitative genetics, population biology, phylogenetics, speciation, and systematics. It has been designed as a companion to Glossary of cellular and molecular biology, which contains many overlapping and related terms; other related glossaries include Glossary of biology and Glossary of ecology.




