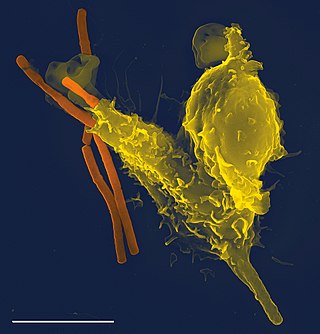
The immune system is a network of biological systems that protects an organism from diseases. It detects and responds to a wide variety of pathogens, from viruses to bacteria, as well as cancer cells, parasitic worms, and also objects such as wood splinters, distinguishing them from the organism's own healthy tissue. Many species have two major subsystems of the immune system. The innate immune system provides a preconfigured response to broad groups of situations and stimuli. The adaptive immune system provides a tailored response to each stimulus by learning to recognize molecules it has previously encountered. Both use molecules and cells to perform their functions.

Toll-like receptors (TLRs) are a class of proteins that play a key role in the innate immune system. They are single-spanning receptors usually expressed on sentinel cells such as macrophages and dendritic cells, that recognize structurally conserved molecules derived from microbes. Once these microbes have reached physical barriers such as the skin or intestinal tract mucosa, they are recognized by TLRs, which activate immune cell responses. The TLRs include TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12, and TLR13. Humans lack genes for TLR11, TLR12 and TLR13 and mice lack a functional gene for TLR10. The receptors TLR1, TLR2, TLR4, TLR5, TLR6, and TLR10 are located on the cell membrane, whereas TLR3, TLR7, TLR8, and TLR9 are located in intracellular vesicles.

Lassa mammarenavirus (LASV) is an arenavirus that causes Lassa hemorrhagic fever, a type of viral hemorrhagic fever (VHF), in humans and other primates. Lassa mammarenavirus is an emerging virus and a select agent, requiring Biosafety Level 4-equivalent containment. It is endemic in West African countries, especially Sierra Leone, the Republic of Guinea, Nigeria, and Liberia, where the annual incidence of infection is between 300,000 and 500,000 cases, resulting in 5,000 deaths per year.
Viral pathogenesis is the study of the process and mechanisms by which viruses cause diseases in their target hosts, often at the cellular or molecular level. It is a specialized field of study in virology.
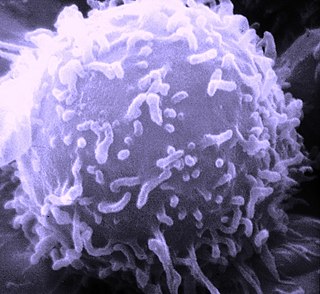
The adaptive immune system, AIS, also known as the acquired immune system, or specific immune system is a subsystem of the immune system that is composed of specialized cells, organs, and processes that eliminate pathogens specifically. The acquired immune system is one of the two main immunity strategies found in vertebrates.
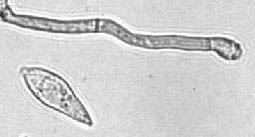
Magnaporthe grisea, also known as rice blast fungus, rice rotten neck, rice seedling blight, blast of rice, oval leaf spot of graminea, pitting disease, ryegrass blast, Johnson spot, neck blast, wheat blast and Imochi (稲熱), is a plant-pathogenic fungus and model organism that causes a serious disease affecting rice. It is now known that M. grisea consists of a cryptic species complex containing at least two biological species that have clear genetic differences and do not interbreed. Complex members isolated from Digitaria have been more narrowly defined as M. grisea. The remaining members of the complex isolated from rice and a variety of other hosts have been renamed Magnaporthe oryzae, within the same M. grisea complex. Confusion on which of these two names to use for the rice blast pathogen remains, as both are now used by different authors.

The innate immune system or nonspecific immune system is one of the two main immunity strategies in vertebrates. The innate immune system is an alternate defense strategy and is the dominant immune system response found in plants, fungi, prokaryotes, and invertebrates.
Systemic acquired resistance (SAR) is a "whole-plant" resistance response that occurs following an earlier localized exposure to a pathogen. SAR is analogous to the innate immune system found in animals, and although there are many shared aspects between the two systems, it is thought to be a result of convergent evolution. The systemic acquired resistance response is dependent on the plant hormone, salicylic acid.
Antigenic variation or antigenic alteration refers to the mechanism by which an infectious agent such as a protozoan, bacterium or virus alters the proteins or carbohydrates on its surface and thus avoids a host immune response, making it one of the mechanisms of antigenic escape. It is related to phase variation. Antigenic variation not only enables the pathogen to avoid the immune response in its current host, but also allows re-infection of previously infected hosts. Immunity to re-infection is based on recognition of the antigens carried by the pathogen, which are "remembered" by the acquired immune response. If the pathogen's dominant antigen can be altered, the pathogen can then evade the host's acquired immune system. Antigenic variation can occur by altering a variety of surface molecules including proteins and carbohydrates. Antigenic variation can result from gene conversion, site-specific DNA inversions, hypermutation, or recombination of sequence cassettes. The result is that even a clonal population of pathogens expresses a heterogeneous phenotype. Many of the proteins known to show antigenic or phase variation are related to virulence.

Hypersensitive response (HR) is a mechanism used by plants to prevent the spread of infection by microbial pathogens. HR is characterized by the rapid death of cells in the local region surrounding an infection and it serves to restrict the growth and spread of pathogens to other parts of the plant. It is analogous to the innate immune system found in animals, and commonly precedes a slower systemic response, which ultimately leads to systemic acquired resistance (SAR). HR can be observed in the vast majority of plant species and is induced by a wide range of plant pathogens such as oomycetes, viruses, fungi and even insects.
The gene-for-gene relationship is a concept in plant pathology that plants and their diseases each have single genes that interact with each other during an infection. It was proposed by Harold Henry Flor who was working with rust (Melampsora lini) of flax (Linum usitatissimum). Flor showed that the inheritance of both resistance in the host and parasite ability to cause disease is controlled by pairs of matching genes. One is a plant gene called the resistance (R) gene. The other is a parasite gene called the avirulence (Avr) gene. Plants producing a specific R gene product are resistant towards a pathogen that produces the corresponding Avr gene product. Gene-for-gene relationships are a widespread and very important aspect of plant disease resistance. Another example can be seen with Lactuca serriola versus Bremia lactucae.
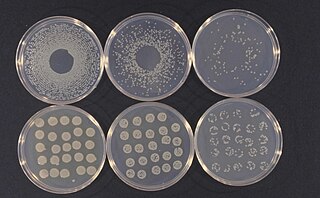
Pseudomonas syringae is a rod-shaped, Gram-negative bacterium with polar flagella. As a plant pathogen, it can infect a wide range of species, and exists as over 50 different pathovars, all of which are available to researchers from international culture collections such as the NCPPB, ICMP, and others.
Plant disease epidemiology is the study of disease in plant populations. Much like diseases of humans and other animals, plant diseases occur due to pathogens such as bacteria, viruses, fungi, oomycetes, nematodes, phytoplasmas, protozoa, and parasitic plants. Plant disease epidemiologists strive for an understanding of the cause and effects of disease and develop strategies to intervene in situations where crop losses may occur. Destructive and non-destructive methods are used to detect diseases in plants. Additionally, understanding the responses of the immune system in plants will further benefit and limit the loss of crops. Typically successful intervention will lead to a low enough level of disease to be acceptable, depending upon the value of the crop.
Resistance genes (R-Genes) are genes in plant genomes that convey plant disease resistance against pathogens by producing R proteins. The main class of R-genes consist of a nucleotide binding domain (NB) and a leucine rich repeat (LRR) domain(s) and are often referred to as (NB-LRR) R-genes or NLRs. Generally, the NB domain binds either ATP/ADP or GTP/GDP. The LRR domain is often involved in protein-protein interactions as well as ligand binding. NB-LRR R-genes can be further subdivided into toll interleukin 1 receptor (TIR-NB-LRR) and coiled-coil (CC-NB-LRR).
Biotic stress is stress that occurs as a result of damage done to an organism by other living organisms, such as bacteria, viruses, fungi, parasites, beneficial and harmful insects, weeds, and cultivated or native plants. It is different from abiotic stress, which is the negative impact of non-living factors on the organisms such as temperature, sunlight, wind, salinity, flooding and drought. The types of biotic stresses imposed on an organism depend the climate where it lives as well as the species' ability to resist particular stresses. Biotic stress remains a broadly defined term and those who study it face many challenges, such as the greater difficulty in controlling biotic stresses in an experimental context compared to abiotic stress.
In plant biology, elicitors are extrinsic or foreign molecules often associated with plant pests, diseases or synergistic organisms. Elicitor molecules can attach to special receptor proteins located on plant cell membranes. These receptors are able to recognize the molecular pattern of elicitors and trigger intracellular defence signalling via the octadecanoid pathway. This response results in the enhanced synthesis of metabolites which reduce damage and increase resistance to pest, disease or environmental stress. This is an immune response called pattern triggered immunity (PTI).
In biology, a pathogen, in the oldest and broadest sense, is any organism or agent that can produce disease. A pathogen may also be referred to as an infectious agent, or simply a germ.

Effector-triggered immunity (ETI) is one of the pathways, along with the pattern-triggered immunity (PTI) pathway, by which the innate immune system recognises pathogenic organisms and elicits a protective immune response. ETI is elicited when an effector protein secreted by a pathogen into the host cell is successfully recognised by the host. Alternatively, effector-triggered susceptibility (ETS) can occur if an effector protein can block the immune response triggered by pattern recognition receptors (PRR) and evade immunity, allowing the pathogen to propagate in the host.

EF-Tu receptor, abbreviated as EFR, is a pattern-recognition receptor (PRR) that binds to the prokaryotic protein EF-Tu in Arabidopsis thaliana. This receptor is an important part of the plant immune system as it allows the plant cells to recognize and bind to EF-Tu, preventing genetic transformation by and protein synthesis in pathogens such as Agrobacterium.

Fungal effectors are proteins or non-proteinaceous molecules secreted by pathogenic fungi into a host organism in order to modulate the host's immune response.



