Related Research Articles

Inbreeding is the production of offspring from the mating or breeding of individuals or organisms that are closely related genetically. By analogy, the term is used in human reproduction, but more commonly refers to the genetic disorders and other consequences that may arise from expression of deleterious recessive traits resulting from incestuous sexual relationships and consanguinity. Animals avoid inbreeding only rarely.
Small populations can behave differently from larger populations. They are often the result of population bottlenecks from larger populations, leading to loss of heterozygosity and reduced genetic diversity and loss or fixation of alleles and shifts in allele frequencies. A small population is then more susceptible to demographic and genetic stochastic events, which can impact the long-term survival of the population. Therefore, small populations are often considered at risk of endangerment or extinction, and are often of conservation concern.

Ex situ conservation is the process of protecting an endangered species, variety, or breed of plant or animal outside its natural habitat. For example, by removing part of the population from a threatened habitat and placing it in a new location, an artificial environment which is similar to the natural habitat of the respective animal and within the care of humans, such as a zoological park or wildlife sanctuary. The degree to which humans control or modify the natural dynamics of the managed population varies widely, and this may include alteration of living environments, reproductive patterns, access to resources, and protection from predation and mortality.

In population genetics, gene flow is the transfer of genetic material from one population to another. If the rate of gene flow is high enough, then two populations will have equivalent allele frequencies and therefore can be considered a single effective population. It has been shown that it takes only "one migrant per generation" to prevent populations from diverging due to drift. Populations can diverge due to selection even when they are exchanging alleles, if the selection pressure is strong enough. Gene flow is an important mechanism for transferring genetic diversity among populations. Migrants change the distribution of genetic diversity among populations, by modifying allele frequencies. High rates of gene flow can reduce the genetic differentiation between the two groups, increasing homogeneity. For this reason, gene flow has been thought to constrain speciation and prevent range expansion by combining the gene pools of the groups, thus preventing the development of differences in genetic variation that would have led to differentiation and adaptation. In some cases dispersal resulting in gene flow may also result in the addition of novel genetic variants under positive selection to the gene pool of a species or population
Heterosis, hybrid vigor, or outbreeding enhancement is the improved or increased function of any biological quality in a hybrid offspring. An offspring is heterotic if its traits are enhanced as a result of mixing the genetic contributions of its parents. The heterotic offspring often has traits that are more than the simple addition of the parents' traits, and can be explained by Mendelian or non-Mendelian inheritance. Typical heterotic/hybrid traits of interest in agriculture are higher yield, quicker maturity, stability, drought tolerance etc.
F1 hybrid (also known as filial 1 hybrid) is the first filial generation of offspring of distinctly different parental types. F1 hybrids are used in genetics, and in selective breeding, where the term F1 crossbreed may be used. The term is sometimes written with a subscript, as F1 hybrid. Subsequent generations are called F2, F3, etc.
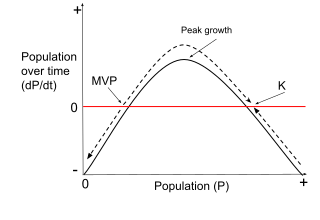
Minimum viable population (MVP) is a lower bound on the population of a species, such that it can survive in the wild. This term is commonly used in the fields of biology, ecology, and conservation biology. MVP refers to the smallest possible size at which a biological population can exist without facing extinction from natural disasters or demographic, environmental, or genetic stochasticity. The term "population" is defined as a group of interbreeding individuals in similar geographic area that undergo negligible gene flow with other groups of the species. Typically, MVP is used to refer to a wild population, but can also be used for ex situ conservation.
Conservation genetics is an interdisciplinary subfield of population genetics that aims to understand the dynamics of genes in a population for the purpose of natural resource management, conservation of genetic diversity, and the prevention of species extinction. Scientists involved in conservation genetics come from a variety of fields including population genetics, research in natural resource management, molecular ecology, molecular biology, evolutionary biology, and systematics. The genetic diversity within species is one of the three fundamental components of biodiversity, so it is an important consideration in the wider field of conservation biology.

Molecular ecology is a subdiscipline of ecology that is concerned with applying molecular genetic techniques to ecological questions. It is virtually synonymous with the field of "Ecological Genetics" as pioneered by Theodosius Dobzhansky, E. B. Ford, Godfrey M. Hewitt, and others. These fields are united in their attempt to study genetic-based questions "out in the field" as opposed to the laboratory. Molecular ecology is related to the fields of population genetics and conservation genetics.
Inbreeding depression is the reduced biological fitness that has the potential to result from inbreeding. The loss of genetic diversity that is seen due to inbreeding, results from small population size. Biological fitness refers to an organism's ability to survive and perpetuate its genetic material. Inbreeding depression is often the result of a population bottleneck. In general, the higher the genetic variation or gene pool within a breeding population, the less likely it is to suffer from inbreeding depression, though inbreeding and outbreeding depression can simultaneously occur.

Captive breeding, also known as captive propagation, is the process of keeping plants or animals in controlled environments, such as wildlife reserves, zoos, botanic gardens, and other conservation facilities. It is sometimes employed to help species that are being threatened by the effects of human activities such as climate change, habitat loss, fragmentation, overhunting or fishing, pollution, predation, disease, and parasitism.
Philopatry is the tendency of an organism to stay in or habitually return to a particular area. The causes of philopatry are numerous, but natal philopatry, where animals return to their birthplace to breed, may be the most common. The term derives from the Greek roots philo, "liking, loving" and patra, "fatherland", although in recent years the term has been applied to more than just the animal's birthplace. Recent usage refers to animals returning to the same area to breed despite not being born there, and migratory species that demonstrate site fidelity: reusing stopovers, staging points, and wintering grounds.
Allogamy or cross-fertilization is the fertilization of an ovum from one individual with the spermatozoa of another. By contrast, autogamy is the term used for self-fertilization. In humans, the fertilization event is an instance of allogamy. Self-fertilization occurs in hermaphroditic organisms where the two gametes fused in fertilization come from the same individual. This is common in plants and certain protozoans.
Extinction vortices are a class of models through which conservation biologists, geneticists and ecologists can understand the dynamics of and categorize extinctions in the context of their causes. This model shows the events that ultimately lead small populations to become increasingly vulnerable as they spiral toward extinction. Developed by M. E. Gilpin and M. E. Soulé in 1986, there are currently four classes of extinction vortices. The first two deal with environmental factors that have an effect on the ecosystem or community level, such as disturbance, pollution, habitat loss etc. Whereas the second two deal with genetic factors such as inbreeding depression and outbreeding depression, genetic drift etc.
Out-crossing or out-breeding is the technique of crossing between different breeds. This is the practice of introducing distantly related genetic material into a breeding line, thereby increasing genetic diversity.
Genetic purging is the increased pressure of natural selection against deleterious alleles prompted by inbreeding.
Inbreeding avoidance, or the inbreeding avoidance hypothesis, is a concept in evolutionary biology that refers to the prevention of the deleterious effects of inbreeding. Animals only rarely exhibit inbreeding avoidance. The inbreeding avoidance hypothesis posits that certain mechanisms develop within a species, or within a given population of a species, as a result of assortative mating and natural and sexual selection, in order to prevent breeding among related individuals. Although inbreeding may impose certain evolutionary costs, inbreeding avoidance, which limits the number of potential mates for a given individual, can inflict opportunity costs. Therefore, a balance exists between inbreeding and inbreeding avoidance. This balance determines whether inbreeding mechanisms develop and the specific nature of such mechanisms.
Autogamy or self-fertilization refers to the fusion of two gametes that come from one individual. Autogamy is predominantly observed in the form of self-pollination, a reproductive mechanism employed by many flowering plants. However, species of protists have also been observed using autogamy as a means of reproduction. Flowering plants engage in autogamy regularly, while the protists that engage in autogamy only do so in stressful environments.

A mixed mating system, also known as “variable inbreeding” a characteristic of many hermaphroditic seed plants, where more than one means of mating is used. Mixed mating usually refers to the production of a mixture of self-fertilized (selfed) and outbred (outcrossed) seeds. Plant mating systems influence the distribution of genetic variation within and among populations, by affecting the propensity of individuals to self-fertilize or cross-fertilize . Mixed mating systems are generally characterized by the frequency of selfing vs. outcrossing, but may include the production of asexual seeds through agamospermy. The trade offs for each strategy depend on ecological conditions, pollinator abundance and herbivory and parasite load. Mating systems are not permanent within species; they can vary with environmental factors, and through domestication when plants are bred for commercial agriculture.

Richard (Dick) Frankham is an Australian biologist, author, and academic. He is an Emeritus Professor in Biology at Macquarie University in Sydney, Australia.
References
 This article incorporates public domain material from Michael Lynch. Inbreeding depression and outbreeding depression. National Park Service.
This article incorporates public domain material from Michael Lynch. Inbreeding depression and outbreeding depression. National Park Service.
- 1 2 3 4 5 6 Frankham, Richard; Ballou, Jonathan D.; Eldridge, Mark D. B.; Lacy, Robert C.; Ralls, Katherine; Dudash, Michele R.; Fenster, Charles B. (2011-04-12). "Predicting the Probability of Outbreeding Depression". Conservation Biology. 25 (3): 465–475. Bibcode:2011ConBi..25..465F. doi: 10.1111/j.1523-1739.2011.01662.x . ISSN 0888-8892. PMID 21486369. S2CID 14824257.
- ↑ Frankham, R.; Ballou, J.D.; Briscoe, D.A. (2002). Introduction to Conservation Genetics . Cambridge. pp. 382. ISBN 0521702712.
- 1 2 3 4 Waser, Nickolas M.; Price, Mary V.; Shaw, Ruth G. (2000). "Outbreeding Depression Varies Among Cohorts of Ipomopsis Aggregata Planted in Nature". Evolution. 54 (2): 485–91. doi:10.1554/0014-3820(2000)054[0485:odvaco]2.0.co;2. ISSN 0014-3820. PMID 10937225.
- ↑ Turcek, FJ (1951). "Effect of introductions on two game populations in Czechoslovakia". Journal of Wildlife Management. 15 (1): 113–114. doi:10.2307/3796784. JSTOR 3796784.
- ↑ Fenster, Charles (2000). "Inbreeding and Outbreeding Depression in Natural Populations of Chamaecrista fasciculata (Fabaceae)". Conservation Biology. 14 (5): 1406–1412. Bibcode:2000ConBi..14.1406F. doi:10.1046/j.1523-1739.2000.99234.x. S2CID 16051555.
- ↑ Erickson and Fenster (2006). "Intraspecific hybridization and the recovery of fitness in the native legume Chamaecrista fasciculata". Evolution. 60 (2): 225–33. doi:10.1554/05-020.1. JSTOR 4095211. PMID 16610315. S2CID 6822055.
- ↑ Lewontin & Birch, R.C. & L.C. (February 3, 1966). "Hybridization as a source of variation for adaptation to new environments". Evolution. 20 (3): 315–336. doi:10.2307/2406633. JSTOR 2406633. PMID 28562982.
- ↑ Frankham, Ballou, & Briscoe, R., J.D. & D.A. (2002). Introduction to Conservation Genetics. Cambridge. p. 388 ISBN 0521702712