
Pterodaustro is a genus of ctenochasmatid pterodactyloid pterosaur from South America. Its fossil remains dated back to the Early Cretaceous period, about 105 million years ago.

Ctenochasma is a genus of Late Jurassic ctenochasmatid pterosaur belonging to the suborder Pterodactyloidea. Three species are currently recognized: C. roemeri, C. taqueti, and C. elegans. Their fossilized remains have been found in the Solnhofen Limestone of Bavaria, Germany, the "Purbeck Group" of northeastern Germany, and the Calcaires tâchetés of eastern France.

Eosipterus is an extinct genus of ctenochasmatid pterosaur from the Early Cretaceous period of Liaoning, China. Fossil remains of Eosipterus dated back to the Aptian stage of the Early Cretaceous, 125 million years ago.
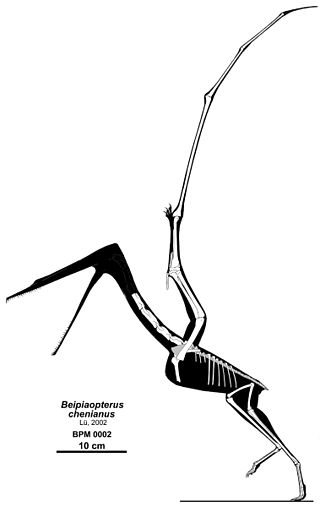
Beipiaopterus is a genus of ctenochasmatid pterosaur from the Early Cretaceous period of what is now the Yixian Formation of the China.
Gegepterus is a genus of ctenochasmatid pterosaur from the Early Cretaceous period of what is now the Yixian Formation of Liaoning, China. Only one species is known, G. changi.
Liaoxipterus is a genus of pterodactyloid pterosaur from the Barremian-Aptian-age Lower Cretaceous Jiufotang Formation of Chaoyang, Liaoning, China. The type species is Liaoxipterus brachyognathus. The genus name is derived from the discovery locality Liaoxi and a Latinised Greek pteron, "wing". The specific name is derived from Greek brachys, "short" and gnathos, "jaw".

Plataleorhynchus is a genus of ctenochasmatid pterodactyloid pterosaur from the Late Jurassic to Early Cretaceous periods of what is now the Purbeck Limestone of Dorset, England.
Liaoningopterus, sometimes misspelled as "Liaoningopteryx", was a genus of anhanguerid pterodactyloid pterosaur from the Barremian-Aptian-age Lower Cretaceous Jiufotang Formation of Chaoyang, Liaoning, China.
Ctenochasmatoidea is a group of early pterosaurs within the suborder Pterodactyloidea. Their remains are usually found in what were once coastal or lake environments. They generally had long wings, long necks, and highly specialized teeth.
Cathayopterus is an extinct genus of ctenochasmatid pterosaur from the Early Cretaceous-age Yixian Formation of Liaoning, China. The name means "China wing", using the word "Cathay" as an old alternative name for China. The type species is C. grabaui, described in 2006 by Wang Xiaolin and Zhou Zhongh. It is a member of the Ctenochasmatidae, a clade of mostly filter feeding pterosaurs from the Jurassic and early Cretaceous.

Ctenochasmatidae is a group of pterosaurs within the suborder Pterodactyloidea. They are characterized by their distinctive teeth, which are thought to have been used for filter-feeding. Ctenochasmatids lived from the Late Jurassic to the Early Cretaceous periods.

Darwinopterus is a genus of pterosaur, discovered in China and named after biologist Charles Darwin. Between 30 and 40 fossil specimens have been identified, all collected from the Tiaojishan Formation, which dates to the middle Jurassic period, 160.89–160.25 Ma ago. The type species, D. modularis, was described in February 2010. D. modularis was the first known pterosaur to display features of both long-tailed (rhamphorhynchoid) and short-tailed (pterodactyloid) pterosaurs, and was described as a transitional fossil between the two groups. Two additional species, D. linglongtaensis and D. robustodens, were described from the same fossil beds in December 2010 and June 2011, respectively.

Euctenochasmatia is an extinct group of pterodactyloid pterosaurs. It was named by David Unwin in 2003 as the group that contains the most recent common ancestor of Pterodactylus and Ctenochasma, and all their descendants.

Gladocephaloideus is a genus of ctenochasmatid ctenochasmatoid pterosaur from the Early Cretaceous period of what is now western Liaoning, China.
Pterofiltrus is a genus of ctenochasmatid pterosaur from the Early Cretaceous of western Liaoning, China.

Moganopterus is an extinct genus of ctenochasmatid pterosaur from the Early Cretaceous of western Liaoning Province, China.
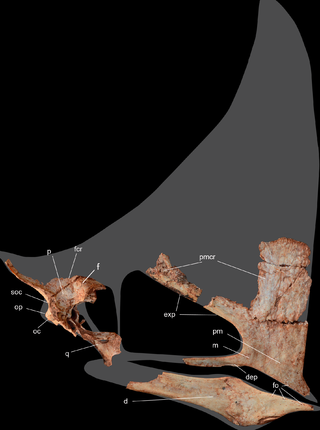
Caiuajara is an extinct genus of tapejarid pterosaur from the Early Cretaceous period of Brazil. It is known from a single type species, Caiuajara dobruskii.
Pangupterus is a genus of pterodactyloid pterosaur from the Early Cretaceous Jiufotang Formation of China. It was first described and named by Lü Junchang et al.

Forfexopterus is a genus of ctenochasmatid pterosaur from the Early Cretaceous Jiufotang Formation in China. It contains a single species, F. jeholensis, named from a mostly complete skeleton by Shunxing Jiang and colleagues in 2016. A second specimen, consisting of a wing, was described in 2020. While the first specimen is larger, it shows signs of being less mature than the second specimen, indicating that the developmental trajectories of Forfexopterus were variable. Like other ctenochasmatids, Forfexopterus had a long, low skull filled with many slender teeth; unlike other members of the group, however, it did not have a spatula-shaped snout tip or crests, and its teeth were more curved. A single characteristic distinguishes Forfexopterus from all other members of the wider group Archaeopterodactyloidea: of the four phalanx bones in its wing finger, the first was shorter than the second but longer than the third.

Caihong is a genus of small paravian theropod dinosaur from China that lived during the Late Jurassic period.














