
The visual system is the physiological basis of visual perception. The system detects, transduces and interprets information concerning light within the visible range to construct an image and build a mental model of the surrounding environment. The visual system is associated with the eye and functionally divided into the optical system and the neural system.

In neuroanatomy, the optic radiation are axons from the neurons in the lateral geniculate nucleus to the primary visual cortex. The optic radiation receives blood through deep branches of the middle cerebral artery and posterior cerebral artery.
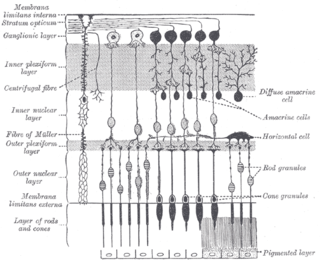
A retinal ganglion cell (RGC) is a type of neuron located near the inner surface of the retina of the eye. It receives visual information from photoreceptors via two intermediate neuron types: bipolar cells and retina amacrine cells. Retina amacrine cells, particularly narrow field cells, are important for creating functional subunits within the ganglion cell layer and making it so that ganglion cells can observe a small dot moving a small distance. Retinal ganglion cells collectively transmit image-forming and non-image forming visual information from the retina in the form of action potential to several regions in the thalamus, hypothalamus, and mesencephalon, or midbrain.
In neuroscience, parvocellular cells, also called P-cells, are neurons located within the parvocellular layers of the lateral geniculate nucleus (LGN) of the thalamus. Their name comes from Latin parvus 'small', due to the small size of the cell compared to the larger magnocellular cells. Phylogenetically, parvocellular neurons are more modern than magnocellular ones.
Magnocellular cells, also called M-cells, are neurons located within the magnocellular layer of the lateral geniculate nucleus of the thalamus. The cells are part of the visual system. They are termed "magnocellular" since they are characterized by their relatively large size compared to parvocellular cells.

In neuroanatomy, the superior colliculus is a structure lying on the roof of the mammalian midbrain. In non-mammalian vertebrates, the homologous structure is known as the optic tectum or optic lobe. The adjective form tectal is commonly used for both structures.

The inferior colliculus (IC) is the principal midbrain nucleus of the auditory pathway and receives input from several peripheral brainstem nuclei in the auditory pathway, as well as inputs from the auditory cortex. The inferior colliculus has three subdivisions: the central nucleus, a dorsal cortex by which it is surrounded, and an external cortex which is located laterally. Its bimodal neurons are implicated in auditory-somatosensory interaction, receiving projections from somatosensory nuclei. This multisensory integration may underlie a filtering of self-effected sounds from vocalization, chewing, or respiration activities.

In neuroanatomy, the pretectal area, or pretectum, is a midbrain structure composed of seven nuclei and comprises part of the subcortical visual system. Through reciprocal bilateral projections from the retina, it is involved primarily in mediating behavioral responses to acute changes in ambient light such as the pupillary light reflex, the optokinetic reflex, and temporary changes to the circadian rhythm. In addition to the pretectum's role in the visual system, the anterior pretectal nucleus has been found to mediate somatosensory and nociceptive information.

In neuroscience, koniocellular cells, also called K-cells, are relatively small neurons located in the koniocellular layer of the lateral geniculate nucleus (LGN) within the thalamus of primates, including humans. The term 'koniocellular' is derived from Greek konio 'dust, poison'.

In neuroanatomy, thalamocortical radiations, also known as thalamocortical fibers, are the efferent fibers that project from the thalamus to distinct areas of the cerebral cortex. They form fiber bundles that emerge from the lateral surface of the thalamus.
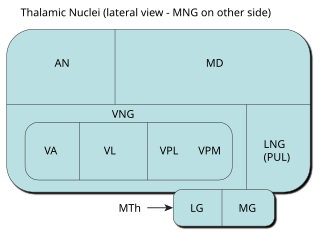
The medial geniculate nucleus (MGN) or medial geniculate body (MGB) is part of the auditory thalamus and represents the thalamic relay between the inferior colliculus (IC) and the auditory cortex (AC). It is made up of a number of sub-nuclei that are distinguished by their neuronal morphology and density, by their afferent and efferent connections, and by the coding properties of their neurons. It is thought that the MGN influences the direction and maintenance of attention.

Retinotopy is the mapping of visual input from the retina to neurons, particularly those neurons within the visual stream. For clarity, 'retinotopy' can be replaced with 'retinal mapping', and 'retinotopic' with 'retinally mapped'.
The zona incerta (ZI) is a horizontally elongated small nucleus that separates the larger subthalamic nucleus from the thalamus. Its connections project extensively over the brain from the cerebral cortex down into the spinal cord.
In neuroanatomy, topographic map is the ordered projection of a sensory surface or an effector system to one or more structures of the central nervous system. Topographic maps can be found in all sensory systems and in many motor systems.
The medial dorsal nucleus is a large nucleus in the thalamus. It is separated from the other thalamic nuclei by the internal medullary lamina.
The isothalamus is a division used by some researchers in describing the thalamus.
Blobs are sections of primary visual cortex (V1) above and below layer IV where groups of neurons sensitive to color assemble in cylindrical shapes. They were first identified in 1979 by Margaret Wong-Riley in cats when she used a cytochrome oxidase stain, from which they get their name. These areas receive input from koniocellular cells in the dorsal lateral geniculate nucleus dLGN and output to the thin stripes of area V2. Interblobs are areas between blobs that receive the same input, but are sensitive to orientation instead of color. They output to the pale and thick stripes of area V2.

A midget cell, sometimes called a P cell or P ganglion cell, is one type of retinal ganglion cell (RGC). Midget cells originate in the ganglion cell layer of the retina, and project to the parvocellular layers of the lateral geniculate nucleus (LGN). The axons of midget cells travel through the optic nerve and optic tract, ultimately synapsing with parvocellular cells in the LGN. These cells are known as midget retinal ganglion cells due to the small sizes of their dendritic trees and cell bodies. About 80% of RGCs are midget cells. They receive inputs from relatively few rods and cones. In many cases, they are connected to midget bipolar cells, which are linked to one cone each.

A parasol cell, sometimes called an M cell or M ganglion cell, is one type of retinal ganglion cell (RGC) located in the ganglion cell layer of the retina. These cells project to magnocellular cells in the lateral geniculate nucleus (LGN) as part of the magnocellular pathway in the visual system. They have large cell bodies as well as extensive branching dendrite networks and as such have large receptive fields. Relative to other RGCs, they have fast conduction velocities. While they do show clear center-surround antagonism, they receive no information about color. Parasol ganglion cells contribute information about the motion and depth of objects to the visual system.

Laura Busse is a German neuroscientist and professor of Systemic Neuroscience within the Division of Neurobiology at the Ludwig Maximilian University of Munich. Busse's lab studies context-dependent visual processing in mouse models by performing large scale in vivo electrophysiological recordings in the thalamic and cortical circuits of awake and behaving mice.










