Related Research Articles
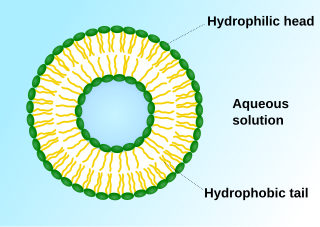
In cell biology, a vesicle is a structure within or outside a cell, consisting of liquid or cytoplasm enclosed by a lipid bilayer. Vesicles form naturally during the processes of secretion (exocytosis), uptake (endocytosis), and the transport of materials within the plasma membrane. Alternatively, they may be prepared artificially, in which case they are called liposomes. If there is only one phospholipid bilayer, the vesicles are called unilamellar liposomes; otherwise they are called multilamellar liposomes. The membrane enclosing the vesicle is also a lamellar phase, similar to that of the plasma membrane, and intracellular vesicles can fuse with the plasma membrane to release their contents outside the cell. Vesicles can also fuse with other organelles within the cell. A vesicle released from the cell is known as an extracellular vesicle.
L1, also known as L1CAM, is a transmembrane protein member of the L1 protein family, encoded by the L1CAM gene. This protein, of 200 to 220 kDa, is a neuronal cell adhesion molecule with a strong implication in cell migration, adhesion, neurite outgrowth, myelination and neuronal differentiation. It also plays a key role in treatment-resistant cancers due to its function. It was first identified in 1984 by M. Schachner who found the protein in post-mitotic mice neurons.
Microparticles are particles between 0.1 and 100 μm in size. Commercially available microparticles are available in a wide variety of materials, including ceramics, glass, polymers, and metals. Microparticles encountered in daily life include pollen, sand, dust, flour, and powdered sugar. The study of microparticles has been called micromeritics, although this term is not very common.

Exosomes, ranging in size from 30 to 150 nanometers, are membrane-bound extracellular vesicles (EVs) that are produced in the endosomal compartment of most eukaryotic cells. In multicellular organisms, exosomes and other EVs are found in biological fluids including saliva, blood, urine and cerebrospinal fluid. EVs have specialized functions in physiological processes, from coagulation and waste management to intercellular communication.

An adrenal tumor or adrenal mass is any benign or malignant neoplasms of the adrenal gland, several of which are notable for their tendency to overproduce endocrine hormones. Adrenal cancer is the presence of malignant adrenal tumors, and includes neuroblastoma, adrenocortical carcinoma and some adrenal pheochromocytomas. Most adrenal pheochromocytomas and all adrenocortical adenomas are benign tumors, which do not metastasize or invade nearby tissues, but may cause significant health problems by unbalancing hormones.

Microvesicles are a type of extracellular vesicle (EV) that are released from the cell membrane. In multicellular organisms, microvesicles and other EVs are found both in tissues and in many types of body fluids. Delimited by a phospholipid bilayer, microvesicles can be as small as the smallest EVs or as large as 1000 nm. They are considered to be larger, on average, than intracellularly-generated EVs known as exosomes. Microvesicles play a role in intercellular communication and can transport molecules such as mRNA, miRNA, and proteins between cells.

CD9 is a gene encoding a protein that is a member of the transmembrane 4 superfamily also known as the tetraspanin family. It is a cell surface glycoprotein that consists of four transmembrane regions and has two extracellular loops that contain disulfide bonds which are conserved throughout the tetraspanin family. Also containing distinct palmitoylation sites that allows CD9 to interact with lipids and other proteins.
Prostasomes are extracellular vesicles secreted by the prostate gland epithelial cells into seminal fluid. They possess an unusual lipid composition and a tight and highly ordered structure of their lipid bilayer membrane, resembling that of lipid raft domains. Prostasomes appear to improve sperm motility and protect against attacks from the female immune defense during the passage to the egg.
ExoCarta is a manually curated database of exosomal proteins, RNA and lipids.
Jan Lötvall is a Swedish clinical allergist and scientist working on translational research primarily in the field of asthma. He is the former director of the Krefting Research Centre at the University of Gothenburg and is the Chief Scientific Officer of ExoCoBio.
Izon Science Limited is a nanotechnology company that develops and sells nano-scale particle analysis and isolation tools. Their main instruments are based on principles of size exclusion chromatography and tunable resistive pulse sensing. Izon’s size-exclusion chromatography columns and related solutions are also used by diagnostics companies focused on developing extracellular vesicle biomarkers.
Extracellular RNA (exRNA) describes RNA species present outside of the cells in which they were transcribed. Carried within extracellular vesicles, lipoproteins, and protein complexes, exRNAs are protected from ubiquitous RNA-degrading enzymes. exRNAs may be found in the environment or, in multicellular organisms, within the tissues or biological fluids such as venous blood, saliva, breast milk, urine, semen, menstrual blood, and vaginal fluid. Although their biological function is not fully understood, exRNAs have been proposed to play a role in a variety of biological processes including syntrophy, intercellular communication, and cell regulation. The United States National Institutes of Health (NIH) published in 2012 a set of Requests for Applications (RFAs) for investigating extracellular RNA biology. Funded by the NIH Common Fund, the resulting program was collectively known as the Extracellular RNA Communication Consortium (ERCC). The ERCC was renewed for a second phase in 2019.
The secretome is the set of proteins expressed by an organism and secreted into the extracellular space. In humans, this subset of the proteome encompasses 13-20% of all proteins, including cytokines, growth factors, extracellular matrix proteins and regulators, and shed receptors. The secretome of a specific tissue can be measured by mass spectrometry and its analysis constitutes a type of proteomics known as secretomics.
The International Society for Extracellular Vesicles (ISEV) is an international scientific organization that focuses on advancing global extracellular vesicle (EV) research. These membrane-bound particles are released from all known cells and include exosomes, ectosomes, exophers, oncosomes, and more. Established in 2011, the society is a nonprofit organization. It is governed by an executive committee. The current president is Kenneth Witwer. Previous presidents were Edit Buzás (2022-2024), Clotilde Théry (2018-2022), Andrew Hill (2016-2018) and founding president Jan Lötvall (2011-2016). The society's journals are the Journal of Extracellular Vesicles and the Journal of Extracellular Biology. ISEV also publishes the international consensus guidelines for EV studies, the "Minimal information for studies of EVs" (MISEV).
The stem cell secretome is a collective term for the paracrine soluble factors produced by stem cells and utilized for their inter-cell communication. In addition to inter-cell communication, the paracrine factors are also responsible for tissue development, homeostasis and (re-)generation. The stem cell secretome consists of extracellular vesicles, specifically exosomes, microvesicles, membrane particles, peptides and small proteins (cytokines). The paracrine activity of stem cells, i.e. the stem cell secretome, has been found to be the predominant mechanism by which stem cell-based therapies mediate their effects in degenerative, auto-immune and/or inflammatory diseases. Though not only stem cells possess a secretome which influences their cellular environment, their secretome currently appears to be the most relevant for therapeutic use.

The Journal of Extracellular Vesicles, JEV, is a peer-reviewed open-access scientific journal of the International Society for Extracellular Vesicles (ISEV). As one of two official journals of ISEV, the other being the Journal of Extracellular Biology, JEV covers research on lipid bilayer-delimited particles known as extracellular vesicles (EVs). EVs are released from cells and include endosome-origin exosomes and plasma membrane-derived ectosomes/microvesicles. The journal was established in 2012 and is currently published by Wiley. The founding editors-in-chief were Clotilde Théry, Yong Song Gho, and Peter Quesenberry. The current editor-in-chief is Jan Lötvall.

Kenneth W. Witwer is an associate professor of molecular and comparative pathobiology and neurology at the Johns Hopkins University School of Medicine in Baltimore, Maryland, United States. He is President of the International Society for Extracellular Vesicles (ISEV) and previously served as Secretary General and Executive Chair of Science and Meetings of the society. His laboratory studies extracellular vesicles (EVs), noncoding and extracellular RNA (exRNA), and enveloped viruses, including HIV and SARS-CoV-2. Witwer is the managing editor of the journal Cytotherapy and a member of the Richman Family Precision Medicine Center of Excellence in Alzheimer's Disease. He has advised the US Environmental Protection Agency and the US National Institutes of Health and is an associate editor of the Journal of Extracellular Vesicles.
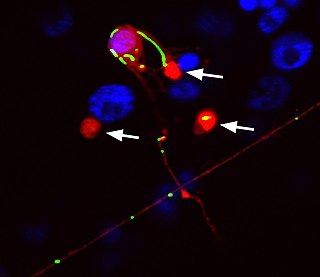
Exophers are a type of membrane-bound extracellular vesicle (EV) that are released by budding out of cells into the extracellular space. Exophers can be released by neurons and muscle in the nematode Caenorhabditis elegans and also from murine cardiomyocytes. Exophers were first discovered in 2017 by an undergraduate student in the lab of Monica Driscoll at Rutgers University.

Intracellular delivery is the process of introducing external materials into living cells. Materials that are delivered into cells include nucleic acids, proteins, peptides, impermeable small molecules, synthetic nanomaterials, organelles, and micron-scale tracers, devices and objects. Such molecules and materials can be used to investigate cellular behavior, engineer cell operations or correct a pathological function.
Exosomes are small vesicles secreted by cells that play a crucial role in intercellular communication. They contain a variety of biomolecules, including proteins, nucleic acids and lipids, which can be transferred between cells to modulate cellular processes. Exosomes have been increasingly acknowledged as promising therapeutic tool and delivery platforms due to unique biological properties.
- Biocompatibility: Exosomes are naturally occurring particles in body, which makes them highly biocompatible and less likely to activate immune response.
- Targeting ability: Exosomes are assembled to express specific proteins or peptides, allowing them to target specific cells or tissues.
- Natural cargo carries: Exosomes can naturally transport a variety of biomolecules, including proteins, RNA and DNA, which can be used for therapeutic purposes.
References
- ↑ Moghassemi, Saeid; Dadashzadeh, Arezoo; Sousa, Maria João; Vlieghe, Hanne; Yang, Jie; León-Félix, Cecibel María; Amorim, Christiani A. (June 2024). "Extracellular vesicles in nanomedicine and regenerative medicine: A review over the last decade". Bioactive Materials. 36: 126–156. doi:10.1016/j.bioactmat.2024.02.021. PMC 10915394 . PMID 38450204.
- ↑ Subedi P, Schneider M, Philipp J, Azimzadeh O, Metzger F, Moertl S, et al. (November 2019). "Comparison of methods to isolate proteins from extracellular vesicles for mass spectrometry-based proteomic analyses". Analytical Biochemistry. 584: 113390. doi: 10.1016/j.ab.2019.113390 . PMID 31401005.
- ↑ Clerici SP, Peppelenbosch M, Fuhler G, Consonni SR, Ferreira-Halder CV (2021-07-15). "Colorectal Cancer Cell-Derived Small Extracellular Vesicles Educate Human Fibroblasts to Stimulate Migratory Capacity". Frontiers in Cell and Developmental Biology. 9: 696373700. doi: 10.3389/fcell.2021.696373 . PMC 8320664 . PMID 34336845.
- ↑ Cordeiro HG, Azevedo-Martins JM, Faria AV, Rocha-Brito KJ, Milani R, Peppelenbosch M, Fuhler G, de Fátima Â, Ferreira-Halder CV (April 2024). "Calix[6]arene dismantles extracellular vesicle biogenesis and metalloproteinases that support pancreatic cancer hallmarks". Cellular Signalling. 119: 111174. doi:10.1016/j.cellsig.2024.111174. PMID 38604340.
- 1 2 3 4 5 6 7 8 Yáñez-Mó M, Siljander PR, Andreu Z, Zavec AB, Borràs FE, Buzas EI, et al. (2015). "Biological properties of extracellular vesicles and their physiological functions". Journal of Extracellular Vesicles. 4: 27066. doi:10.3402/jev.v4.27066. PMC 4433489 . PMID 25979354.
- ↑ Chargaff E, West R (November 1946). "The biological significance of the thromboplastic protein of blood". The Journal of Biological Chemistry. 166 (1): 189–97. doi: 10.1016/S0021-9258(17)34997-9 . PMID 20273687.
- ↑ Wolf P (May 1967). "The nature and significance of platelet products in human plasma". British Journal of Haematology. 13 (3): 269–88. doi:10.1111/j.1365-2141.1967.tb08741.x. PMID 6025241. S2CID 19215210.
- ↑ Anderson HC (April 1969). "Vesicles associated with calcification in the matrix of epiphyseal cartilage". The Journal of Cell Biology. 41 (1): 59–72. doi:10.1083/jcb.41.1.59. PMC 2107736 . PMID 5775794.
- 1 2 Bonucci E (1970). "Fine structure and histochemistry of "calcifying globules" in epiphyseal cartilage". Zeitschrift für Zellforschung und Mikroskopische Anatomie. 103 (2): 192–217. doi:10.1007/BF00337312. PMID 5412827. S2CID 8633696.
- ↑ Nunez EA, Wallis J, Gershon MD (October 1974). "Secretory processes in follicular cells of the bat thyroid. 3. The occurrence of extracellular vesicles and colloid droplets during arousal from hibernation". The American Journal of Anatomy. 141 (2): 179–201. doi:10.1002/aja.1001410203. PMID 4415703.
- ↑ Chandler RL, Bird RG, Bland AP (November 1975). "Letter: Particles associated with microvillous border of intestinal mucosa". Lancet. 2 (7941): 931–2. doi:10.1016/s0140-6736(75)92175-3. PMID 53415. S2CID 40320534.
- ↑ De Broe M, Wieme R, Roels F (December 1975). "Letter: Membrane fragments with koinozymic properties released from villous adenoma of the rectum". Lancet. 2 (7946): 1214–5. doi:10.1016/s0140-6736(75)92709-9. PMID 53703. S2CID 32026872.
- 1 2 Benz EW, Moses HL (June 1974). "Small, virus-like particles detected in bovine sera by electron microscopy". Journal of the National Cancer Institute. 52 (6): 1931–4. doi:10.1093/jnci/52.6.1931. PMID 4834422.
- 1 2 3 Dalton AJ (May 1975). "Microvesicles and vesicles of multivesicular bodies versus "virus-like" particles". Journal of the National Cancer Institute. 54 (5): 1137–48. doi:10.1093/jnci/54.5.1137. PMID 165305.
- ↑ Yim K, Borgoni S, Chahwan R (April 2022). "Letter: Serum extracellular vesicles profiling is associated with COVID-19 progression and immune responses". J Extracell Biol. 1 (4): e37. doi:10.1002/jex2.37. PMC 9088353 . PMID 35574251.
- ↑ Pan BT, Johnstone RM (July 1983). "Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor". Cell. 33 (3): 967–78. doi:10.1016/0092-8674(83)90040-5. PMID 6307529. S2CID 33216388.
- ↑ Harding C, Heuser J, Stahl P (November 1984). "Endocytosis and intracellular processing of transferrin and colloidal gold-transferrin in rat reticulocytes: demonstration of a pathway for receptor shedding". European Journal of Cell Biology. 35 (2): 256–63. PMID 6151502.
- ↑ Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C (July 1987). "Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes)". The Journal of Biological Chemistry. 262 (19): 9412–20. doi: 10.1016/S0021-9258(18)48095-7 . PMID 3597417.
- ↑ Dvorak HF, Quay SC, Orenstein NS, Dvorak AM, Hahn P, Bitzer AM, Carvalho AC (May 1981). "Tumor shedding and coagulation". Science. 212 (4497): 923–4. Bibcode:1981Sci...212..923D. doi:10.1126/science.7195067. PMID 7195067.
- ↑ Fox AS, Yoon SB (November 1970). "DNA-induced transformation in Drosophila: locus-specificity and the establishment of transformed stocks". Proceedings of the National Academy of Sciences of the United States of America. 67 (3): 1608–15. Bibcode:1970PNAS...67.1608F. doi: 10.1073/pnas.67.3.1608 . PMC 283397 . PMID 5274483.
- ↑ Mishra NC, Tatum EL (December 1973). "Non-Mendelian inheritance of DNA-induced inositol independence in Neurospora". Proceedings of the National Academy of Sciences of the United States of America. 70 (12): 3875–9. Bibcode:1973PNAS...70.3875M. doi: 10.1073/pnas.70.12.3875 . PMC 427348 . PMID 4521213.
- ↑ Stegmayr B, Ronquist G (1982). "Promotive effect on human sperm progressive motility by prostasomes". Urological Research. 10 (5): 253–7. doi:10.1007/bf00255932. PMID 6219486. S2CID 26574697.
- ↑ Raposo G, Nijman HW, Stoorvogel W, Liejendekker R, Harding CV, Melief CJ, Geuze HJ (March 1996). "B lymphocytes secrete antigen-presenting vesicles". The Journal of Experimental Medicine. 183 (3): 1161–72. doi:10.1084/jem.183.3.1161. PMC 2192324 . PMID 8642258.
- ↑ Mack M, Kleinschmidt A, Brühl H, Klier C, Nelson PJ, Cihak J, et al. (July 2000). "Transfer of the chemokine receptor CCR5 between cells by membrane-derived microparticles: a mechanism for cellular human immunodeficiency virus 1 infection". Nature Medicine. 6 (7): 769–75. doi:10.1038/77498. PMID 10888925. S2CID 23027144.
- ↑ Rozmyslowicz T, Majka M, Kijowski J, Murphy SL, Conover DO, Poncz M, et al. (January 2003). "Platelet- and megakaryocyte-derived microparticles transfer CXCR4 receptor to CXCR4-null cells and make them susceptible to infection by X4-HIV". AIDS. 17 (1): 33–42. doi: 10.1097/00002030-200301030-00006 . PMID 12478067. S2CID 6619801.
- ↑ Baj-Krzyworzeka M, Szatanek R, Weglarczyk K, Baran J, Urbanowicz B, Brański P, et al. (July 2006). "Tumour-derived microvesicles carry several surface determinants and mRNA of tumour cells and transfer some of these determinants to monocytes". Cancer Immunology, Immunotherapy. 55 (7): 808–18. doi:10.1007/s00262-005-0075-9. PMC 11030663 . PMID 16283305. S2CID 25723677.
- ↑ Ratajczak J, Wysoczynski M, Hayek F, Janowska-Wieczorek A, Ratajczak MZ (September 2006). "Membrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communication". Leukemia. 20 (9): 1487–95. doi:10.1038/sj.leu.2404296. PMID 16791265. S2CID 6874345.
- ↑ Aliotta JM, Sanchez-Guijo FM, Dooner GJ, Johnson KW, Dooner MS, Greer KA, et al. (September 2007). "Alteration of marrow cell gene expression, protein production, and engraftment into lung by lung-derived microvesicles: a novel mechanism for phenotype modulation". Stem Cells. 25 (9): 2245–56. doi:10.1634/stemcells.2007-0128. PMC 3376082 . PMID 17556595.
- ↑ Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO (June 2007). "Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells". Nature Cell Biology. 9 (6): 654–9. doi:10.1038/ncb1596. PMID 17486113. S2CID 8599814.
- ↑ Skog J, Würdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, et al. (December 2008). "Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers". Nature Cell Biology. 10 (12): 1470–6. doi:10.1038/ncb1800. PMC 3423894 . PMID 19011622.
- ↑ Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, van Eijndhoven MA, Hopmans ES, Lindenberg JL, et al. (April 2010). "Functional delivery of viral miRNAs via exosomes". Proceedings of the National Academy of Sciences of the United States of America. 107 (14): 6328–33. Bibcode:2010PNAS..107.6328P. doi: 10.1073/pnas.0914843107 . PMC 2851954 . PMID 20304794.
- 1 2 Chetty VK, Ghanam J, Anchan S, Reinhardt K, Brenzel A, Gelléri M, Cremer C, Grueso-Navarro E, Schneider M, von Neuhoff N, Reinhardt D, Jablonska J, Nazarenko I, Thakur BK (April 2022). "Efficient Small Extracellular Vesicles (EV) Isolation Method and Evaluation of EV-Associated DNA Role in Cell-Cell Communication in Cancer". Cancers (Basel). 14 (9): 2068. doi: 10.3390/cancers14092068 . PMC 9099953 . PMID 35565197.
- ↑ Al-Nedawi K, Meehan B, Rak J (July 2009). "Microvesicles: messengers and mediators of tumor progression". Cell Cycle. 8 (13): 2014–8. doi: 10.4161/cc.8.13.8988 . PMID 19535896.
- ↑ Hoshino A, Costa-Silva B, Shen TL, Rodrigues G, Hashimoto A, Tesic Mark M, et al. (November 2015). "Tumour exosome integrins determine organotropic metastasis". Nature. 527 (7578): 329–35. Bibcode:2015Natur.527..329H. doi:10.1038/nature15756. PMC 4788391 . PMID 26524530.
- ↑ Peinado H, Alečković M, Lavotshkin S, Matei I, Costa-Silva B, Moreno-Bueno G, et al. (June 2012). "Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET". Nature Medicine. 18 (6): 883–91. doi:10.1038/nm.2753. PMC 3645291 . PMID 22635005.
- ↑ Melo SA, Sugimoto H, O'Connell JT, Kato N, Villanueva A, Vidal A, et al. (November 2014). "Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis". Cancer Cell. 26 (5): 707–21. doi:10.1016/j.ccell.2014.09.005. PMC 4254633 . PMID 25446899.
- ↑ Kamerkar S, LeBleu VS, Sugimoto H, Yang S, Ruivo CF, Melo SA, et al. (June 2017). "Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer". Nature. 546 (7659): 498–503. Bibcode:2017Natur.546..498K. doi:10.1038/nature22341. PMC 5538883 . PMID 28607485.
- ↑ Ostrowski M, Carmo NB, Krumeich S, Fanget I, Raposo G, Savina A, et al. (January 2010). "Rab27a and Rab27b control different steps of the exosome secretion pathway". Nature Cell Biology. 12 (1): 19–30, sup pp 1–13. doi:10.1038/ncb2000. hdl: 10044/1/19574 . PMID 19966785. S2CID 13935708.
- ↑ van Niel G, Porto-Carreiro I, Simoes S, Raposo G (July 2006). "Exosomes: a common pathway for a specialized function". Journal of Biochemistry. 140 (1): 13–21. doi: 10.1093/jb/mvj128 . PMID 16877764. S2CID 43541754.
- ↑ Kowal J, Arras G, Colombo M, Jouve M, Morath JP, Primdal-Bengtson B, et al. (February 2016). "Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes". Proceedings of the National Academy of Sciences of the United States of America. 113 (8): E968-77. Bibcode:2016PNAS..113E.968K. doi: 10.1073/pnas.1521230113 . PMC 4776515 . PMID 26858453.
- ↑ Tkach M, Kowal J, Théry C (January 2018). "Why the need and how to approach the functional diversity of extracellular vesicles". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 373 (1737): 20160479. doi:10.1098/rstb.2016.0479. PMC 5717434 . PMID 29158309.
- ↑ Leslie M (August 2013). "Cell Biology. NIH effort gambles on mysterious extracellular RNAs". Science. 341 (6149): 947. doi:10.1126/science.341.6149.947. PMID 23990535.
- 1 2 Gutierrez BC, Ancarola ME, Volpato-Rossi I, Marcilla A, Ramirez MI, Rosenzvit MC, et al. (2022). "Extracellular vesicles from Trypanosoma cruzi-dendritic cell interaction show modulatory properties and confer resistance to lethal infection as a cell-free based therapy strategy". Frontiers in Cellular and Infection Microbiology. 12: 980817. doi: 10.3389/fcimb.2022.980817 . PMC 9710384 . PMID 36467728.
- ↑ Bazzan E, Tinè M, Casara A, Biondini D, Semenzato U, Cocconcelli E, et al. (June 2021). "Critical Review of the Evolution of Extracellular Vesicles' Knowledge: From 1946 to Today". International Journal of Molecular Sciences. 22 (12): 6417. doi: 10.3390/ijms22126417 . PMC 8232679 . PMID 34203956.
- ↑ Gurunathan S, Kang MH, Jeyaraj M, Qasim M, Kim JH (April 2019). "Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes". Cells. 8 (4): 307. doi: 10.3390/cells8040307 . PMC 6523673 . PMID 30987213.
- 1 2 3 4 5 6 7 Théry C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, et al. (2018). "Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines". Journal of Extracellular Vesicles. 7 (1): 1535750. doi:10.1080/20013078.2018.1535750. PMC 6322352 . PMID 30637094.
- ↑ Trams EG, Lauter CJ, Salem N, Heine U (July 1981). "Exfoliation of membrane ecto-enzymes in the form of micro-vesicles". Biochimica et Biophysica Acta (BBA) - Biomembranes. 645 (1): 63–70. doi:10.1016/0005-2736(81)90512-5. PMID 6266476.
- ↑ Morello M, Minciacchi VR, de Candia P, Yang J, Posadas E, Kim H, et al. (November 2013). "Large oncosomes mediate intercellular transfer of functional microRNA". Cell Cycle. 12 (22): 3526–36. doi:10.4161/cc.26539. PMC 3906338 . PMID 24091630.
- ↑ Meehan B, Rak J, Di Vizio D (2016). "Oncosomes - large and small: what are they, where they came from?". Journal of Extracellular Vesicles. 5: 33109. doi:10.3402/jev.v5.33109. PMC 5040817 . PMID 27680302.
- ↑ Minciacchi VR, Spinelli C, Reis-Sobreiro M, Cavallini L, You S, Zandian M, Li X, Mishra R, Chiarugi P, Adam RM, Posadas EM, Viglietto G, Freeman MR, Cocucci E, Bhowmick NA, Di Vizio D (2017). "MYC Mediates Large Oncosome-Induced Fibroblast Reprogramming in Prostate Cancer". Cancer Research. 77 (9): 2306–2317. doi: 10.1158/0008-5472.CAN-16-2942 . PMID 28202510.
- ↑ Bertolini I, Terrasi A, Martelli C, Gaudioso G, Di Cristofori A, Storaci AM, Formica M, Braidotti P, Todoerti K, Ferrero S, Caroli M, Ottobrini L, Vaccari T, Vaira V (2019). "A GBM-like V-ATPase signature directs cell-cell tumor signaling and reprogramming via large oncosomes". eBioMedicine. 41: 225–235. doi:10.1016/j.ebiom.2019.01.051. PMC 6441844 . PMID 30737083.
- 1 2 3 Melentijevic I, Toth ML, Arnold ML, Guasp RJ, Harinath G, Nguyen KC, et al. (February 2017). "C. elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress". Nature. 542 (7641): 367–371. Bibcode:2017Natur.542..367M. doi:10.1038/nature21362. PMC 5336134 . PMID 28178240.
- ↑ Nicolás-Ávila JA, Lechuga-Vieco AV, Esteban-Martínez L, Sánchez-Díaz M, Díaz-García E, Santiago DJ, et al. (2020). "A Network of Macrophages Supports Mitochondrial Homeostasis in the Heart". Cell. 183 (1): 94–109. doi: 10.1016/j.cell.2020.08.031 . hdl: 10261/226682 . PMID 32937105. S2CID 221716195.
- ↑ Ma L, Li Y, Peng J, Wu D, Zhao X, Cui Y, Chen L, Yan X, Du Y, Yu L (2015). "Discovery of the migrasome, an organelle mediating release of cytoplasmic contents during cell migration". Cell Research. 25 (1): 24–38. doi:10.1038/cr.2014.135. PMC 4650581 . PMID 25342562.
- ↑ Jiao H, Jiang D, Hu X, Du W, Ji L, Yang Y, Li X, Sho T, Wang X, Li Y, Wu YT, Wei YH, Hu X, Yu L (2021). "Mitocytosis, a migrasome-mediated mitochondrial quality-control process". Cell. 184 (11): 2896–2910. doi: 10.1016/j.cell.2021.04.027 . PMID 34048705. S2CID 235226529.
- ↑ Nolte-'t Hoen E, Cremer T, Gallo RC, Margolis LB (August 2016). "Extracellular vesicles and viruses: Are they close relatives?". Proceedings of the National Academy of Sciences of the United States of America. 113 (33): 9155–61. Bibcode:2016PNAS..113.9155N. doi: 10.1073/pnas.1605146113 . PMC 4995926 . PMID 27432966.
- ↑ Mateescu B, Kowal EJ, van Balkom BW, Bartel S, Bhattacharyya SN, Buzás EI, et al. (2017). "Obstacles and opportunities in the functional analysis of extracellular vesicle RNA - an ISEV position paper". Journal of Extracellular Vesicles. 6 (1): 1286095. doi:10.1080/20013078.2017.1286095. PMC 5345583 . PMID 28326170.
- ↑ Multia E, Tear CJ, Palviainen M, Siljander P, Riekkola ML (December 2019). "Fast isolation of highly specific population of platelet-derived extracellular vesicles from blood plasma by affinity monolithic column, immobilized with anti-human CD61 antibody". Analytica Chimica Acta. 1091: 160–168. Bibcode:2019AcAC.1091..160M. doi:10.1016/j.aca.2019.09.022. hdl: 10138/321264 . PMID 31679569. S2CID 203147714.
- ↑ Multia E, Liangsupree T, Jussila M, Ruiz-Jimenez J, Kemell M, Riekkola ML (October 2020). "Automated On-Line Isolation and Fractionation System for Nanosized Biomacromolecules from Human Plasma". Analytical Chemistry. 92 (19): 13058–13065. doi:10.1021/acs.analchem.0c01986. PMC 7586295 . PMID 32893620.
- ↑ Morani M, Mai TD, Krupova Z, Defrenaix P, Multia E, Riekkola ML, Taverna M (September 2020). "Electrokinetic characterization of extracellular vesicles with capillary electrophoresis: A new tool for their identification and quantification". Analytica Chimica Acta. 1128: 42–51. Bibcode:2020AcAC.1128...42M. doi:10.1016/j.aca.2020.06.073. hdl: 10138/332354 . PMID 32825911. S2CID 221238347.
- ↑ Lacroix R, Judicone C, Poncelet P, Robert S, Arnaud L, Sampol J, Dignat-George F (March 2012). "Impact of pre-analytical parameters on the measurement of circulating microparticles: towards standardization of protocol". Journal of Thrombosis and Haemostasis. 10 (3): 437–46. doi: 10.1111/j.1538-7836.2011.04610.x . PMID 22212198. S2CID 46519893.
- ↑ Witwer KW, Buzás EI, Bemis LT, Bora A, Lässer C, Lötvall J, et al. (2013). "Standardization of sample collection, isolation and analysis methods in extracellular vesicle research". Journal of Extracellular Vesicles. 2: 20360. doi:10.3402/jev.v2i0.20360. PMC 3760646 . PMID 24009894.
- ↑ Coumans FA, Brisson AR, Buzas EI, Dignat-George F, Drees EE, El-Andaloussi S, et al. (May 2017). "Methodological Guidelines to Study Extracellular Vesicles". Circulation Research. 120 (10): 1632–1648. doi: 10.1161/CIRCRESAHA.117.309417 . PMID 28495994.
- ↑ Liangsupree T, Multia E, Riekkola ML (January 2021). "Modern isolation and separation techniques for extracellular vesicles". Journal of Chromatography A. 1636: 461773. doi: 10.1016/j.chroma.2020.461773 . ISSN 0021-9673. PMID 33316564.
- ↑ Lötvall J, Hill AF, Hochberg F, Buzás EI, Di Vizio D, Gardiner C, et al. (2014). "Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the International Society for Extracellular Vesicles". Journal of Extracellular Vesicles. 3: 26913. doi:10.3402/jev.v3.26913. PMC 4275645 . PMID 25536934.
- ↑ Van Deun J, Mestdagh P, Agostinis P, Akay Ö, Anand S, Anckaert J, et al. (February 2017). "EV-TRACK: transparent reporting and centralizing knowledge in extracellular vesicle research". Nature Methods. 14 (3): 228–232. doi:10.1038/nmeth.4185. PMID 28245209. S2CID 205425936.
- ↑ Dhondt B, Geeurickx E, Tulkens J, Van Deun J, Vergauwen G, Lippens L, et al. (11 March 2020). "Unravelling the proteomic landscape of extracellular vesicles in prostate cancer by density-based fractionation of urine". Journal of Extracellular Vesicles. 9 (1): 1736935. doi: 10.1080/20013078.2020.1736935 . PMC 7144211 . PMID 32284825.
- ↑ Yin Y, Chen H, Wang Y, Zhang L, Wang X (October 2021). "Roles of extracellular vesicles in the aging microenvironment and age-related diseases". Journal of Extracellular Vesicles. 10 (12): e12154. doi:10.1002/jev2.12154. PMC 8491204 . PMID 34609061.
- ↑ Xu D, Tahara H (March 2013). "The role of exosomes and microRNAs in senescence and aging". Advanced Drug Delivery Reviews. 65 (3): 368–375. doi:10.1016/j.addr.2012.07.010. PMID 22820533.
- ↑ Suh N (October 2018). "MicroRNA controls of cellular senescence". BMB Reports. 51 (10): 493–499. doi:10.5483/BMBRep.2018.51.10.209. PMC 6235093 . PMID 30269742.
- ↑ Jiang F, Chen Q, Wang W, Ling Y, Yan Y, Xia P (January 2020). "Hepatocyte-derived extracellular vesicles promote endothelial inflammation and atherogenesis via microRNA-1". Journal of Hepatology. 72 (1): 156–166. doi:10.1016/j.jhep.2019.09.014. PMID 31568800. S2CID 203622470.
- ↑ Grigorian Shamagian L, Rogers RG, Luther K, Angert D, Echavez A, Liu W, et al. (July 2023). "Rejuvenating effects of young extracellular vesicles in aged rats and in cellular models of human senescence". Scientific Reports. 13 (1): 12240. Bibcode:2023NatSR..1312240G. doi:10.1038/s41598-023-39370-5. PMC 10382547 . PMID 37507448.
- ↑ Owens, A. Phillip; Mackman, Nigel (2011-05-13). Weber, Christian; Mause, Sebastian (eds.). "Microparticles in Hemostasis and Thrombosis". Circulation Research. 108 (10): 1284–1297. doi:10.1161/CIRCRESAHA.110.233056. ISSN 0009-7330. PMC 3144708 . PMID 21566224.
- ↑ Yamamoto S, Azuma E, Muramatsu M, Hamashima T, Ishii Y, Sasahara M (November 2016). "Significance of Extracellular Vesicles: Pathobiological Roles in Disease". Cell Structure and Function. 41 (2): 137–143. doi: 10.1247/csf.16014 . PMID 27679938.
- ↑ Yim K, AlHrout A, Borgoni S, Chahwan R (December 2020). "Extracellular Vesicles Orchestrate Immune and Tumor Interaction Networks". Cancers. 12 (12): 3696. doi: 10.3390/cancers12123696 . PMC 7763968 . PMID 33317058.
- ↑ Cappariello A, Rucci N (September 2019). "Tumour-Derived Extracellular Vesicles (EVs): A Dangerous "Message in A Bottle" for Bone". International Journal of Molecular Sciences. 20 (19): 4805. doi: 10.3390/ijms20194805 . PMC 6802008 . PMID 31569680.
- ↑ Makhijani, Priya; McGaha, Tracy L. (2022). "Myeloid Responses to Extracellular Vesicles in Health and Disease". Frontiers in Immunology. 13: 818538. doi: 10.3389/fimmu.2022.818538 . ISSN 1664-3224. PMC 8934876 . PMID 35320943.
- ↑ Söllvander S, Nikitidou E, Brolin R, Söderberg L, Sehlin D, Lannfelt L, Erlandsson A (May 2016). "Accumulation of amyloid-β by astrocytes result in enlarged endosomes and microvesicle-induced apoptosis of neurons". Molecular Neurodegeneration. 11 (1): 38. doi: 10.1186/s13024-016-0098-z . PMC 4865996 . PMID 27176225.
- ↑ Nikitidou E, Khoonsari PE, Shevchenko G, Ingelsson M, Kultima K, Erlandsson A (2017). "Increased Release of Apolipoprotein E in Extracellular Vesicles Following Amyloid-β Protofibril Exposure of Neuroglial Co-Cultures". Journal of Alzheimer's Disease. 60 (1): 305–321. doi:10.3233/JAD-170278. PMC 5676865 . PMID 28826183.
- ↑ Georgievski A, Michel A, Thomas C, Mlamla Z, Pais de Barros JP, Lemaire-Ewing S, et al. (2022). "Acute lymphoblastic leukemia-derived extracellular vesicles affect quiescence of hematopoietic stem and progenitor cells". Cell Death Dis. 12 (4): 337. doi:10.1038/s41419-022-04761-5. PMC 9005650 . PMID 35414137.
- ↑ Lanna A, Vaz B, D'Ambra C, Valvo S, Vuotto C, Chiurchiù V, et al. (October 2022). "An intercellular transfer of telomeres rescues T cells from senescence and promotes long-term immunological memory". Nature Cell Biology. 24 (10): 1461–1474. doi:10.1038/s41556-022-00991-z. PMC 7613731 . PMID 36109671.
- ↑ Agnati LF, Guidolin D, Guescini M, Genedani S, Fuxe K (September 2010). "Understanding wiring and volume transmission". Brain Research Reviews. 64 (1): 137–159. doi:10.1016/j.brainresrev.2010.03.003. PMID 20347870. S2CID 36665895.
- ↑ Koniusz S, Andrzejewska A, Muraca M, Srivastava AK, Janowski M, Lukomska B (2016). "Extracellular Vesicles in Physiology, Pathology, and Therapy of the Immune and Central Nervous System, with Focus on Extracellular Vesicles Derived from Mesenchymal Stem Cells as Therapeutic Tools". Frontiers in Cellular Neuroscience. 10: 109. doi: 10.3389/fncel.2016.00109 . PMC 4852177 . PMID 27199663.
- ↑ Verderio C, Muzio L, Turola E, Bergami A, Novellino L, Ruffini F, et al. (October 2012). "Myeloid microvesicles are a marker and therapeutic target for neuroinflammation". Annals of Neurology. 72 (4): 610–624. doi:10.1002/ana.23627. PMID 23109155. S2CID 35702508.
- ↑ Urbanelli L, Buratta S, Sagini K, Ferrara G, Lanni M, Emiliani C (2015). "Exosome-based strategies for Diagnosis and Therapy". Recent Patents on CNS Drug Discovery. 10 (1): 10–27. doi:10.2174/1574889810666150702124059. PMID 26133463.
- ↑ Skog J, Würdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, et al. (December 2008). "Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers". Nature Cell Biology. 10 (12): 1470–1476. doi:10.1038/ncb1800. PMC 3423894 . PMID 19011622.