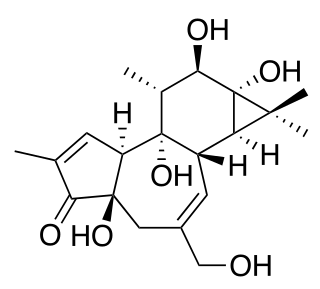
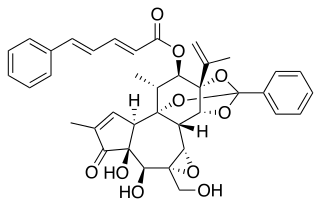
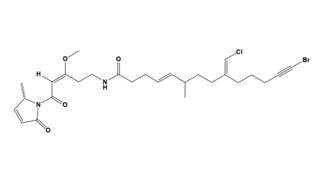
Debromoaplysiatoxin is a toxic agent produced by the blue-green alga Lyngbya majuscula. This alga lives in marine waters and causes seaweed dermatitis. Furthermore, it is a tumor promoter which has an anti-proliferative activity against various cancer cell lines in mice.
The first reported case of seaweed dermatitis was from 1958 in Hawaii on Oahu island. About 125 people who had been swimming in the sea get suffered from symptoms like itching, burning, blisters, rash and desquamation. The causative substance of this seaweed dermatitis was not known till 1968 when people in Okinawa, Japan, suffered from the same symptoms as the people in Hawaii. After researchers took samples in 1973 from Lyngbya majuscula they found out that this was the causative agent of the dermatitis.[1][2]
In 1980 there was a new outbreak of seaweed dermatitis on Oahu island Hawaii. Samples of L. majuscula revealed that this blue-green alga contained a mixture of aplysiatoxin, debromoaplysiatoxin and lyngbyatoxin A. These three substances appeared to be the causative toxins of seaweed dermatitis.[2]
Years later, in 1994, local people of Hawaii, Maui and Oahu island in Hawaii were poisoned by food. The local residents of these islands often eat various types of algae including the red alga Gracilaria coronopifolia. After taking samples of this red alga it turned out that they contained two toxins which were identical with aplysiatoxin and debromoaplysiatoxin. Moreover, they observed parasitism of a blue-green alga on the surface of G. coronopifolia. In view of the fact that some blue-green alga like L. majuscula produce aplysiatoxin and debromoaplysiatoxin, it's probable that they are the true origin of this food poisoning case.[3]
Synthesis
Because of the sterically complex and particular molecular structure, debromoaplysiatoxin is an attractive target for total synthesis. This compound combines spiro acetal, hemiacetal and diolide functionalities, which result in peculiar biological activities.[4] To this date, only Yoshita Kishi's approach to synthesize debromoaplysiatoxin from scratch was found to be effective.[5]
At first, a specific sulfone is formed in 22 steps of common chemistry. This sulfone is coupled with a straightforward epoxide that originates from optically active starting materials.[5] This coupling reaction is most efficient when the sulfone is transformed into a dianion with n-butyllithium, followed by exposure to the epoxide. Hereby, a diastereomeric mixture of sulfones is formed. After reductive desulfurization and methylation, the formation of a cyclohexylidene side group can be achieved. This intermediate can be converted into a terminal epoxide by usage of classical synthetical operations, such as introducing acetic acid or substitution reactions with tosylates. Next, a dithiane derived anion interacts to form an alcohol.
The following step involves introducing an acidic sidechain on the C.9 hydroxyl group. This carboxylic, acidic side group is made of xylose and arabidose. The introduction is realized by protecting the C.29 and C.30 hydroxylgroups with respectively p-methoxybenzyl (MPM) and benzyloxymethyl (BOM), activating the sidechain with acid chloride, and subsequently replacing the C.30 protecting group with a more stable one, tert-butyldiphenylsilyl (TBDPS). Thereafter, the carboxylic acid can successfully attach to the backbone by esterification. In order to produce the primary alcohol, it is necessary to adjust the protecting group of the C.30 once again, back to BOM.
In order to introduce the ring system that characterizes debromoaplysiotoxin in this molecule, hemiketal formation must be achieved between the C.7 ketone and the C.11 hydroxygroup, after which lactone formation can seal the deal. This can be achieved by a method called macrolactonization.
Mechanism of action
Debromoaplysiatoxin has two effects on cell-growth: it has a tumor-promoting-activity as well as an anti-proliferation activity. Therefore, this compound is of interest for study: by finding which groups cause the tumor-promoting activity, and removing them, the new compound will only have an anti-proliferation activity, and can be used as a therapy for cancer. The methoxy-group is found to be a cause for the tumor-promoting activity, and removal of the methoxy-group causes the antiproliferation activity to increase, without changing the tumor-promoting activity. Studies with analogs of debromoaplysiatoxin show that the hemi-acetalhydroxy group at position 3 and/or the methoxy group at position 15 in debromoaplysiatoxin would be responsible for the tumor-promoting activity.
The activation of protein kinase C (PKC) is likely to be responsible for the antiproliferative and tumor-promoting activities of aplysiatoxin-related compounds. Tumor promoters like PDBu and ATX bound potently to the C1 domains of both conventional and novel PKCs, while antiproliferative compounds such as aplog-1 and bryo-1 exhibited some selectivity for novel PKCs other than PKCε, that is, PKCδ, PKCη, and PKCθ. The affinity of DAT for the PKC C1 domains is quite similar to that of aplysiatoxin. The introduction of a methylgroup group into position 4 could enhance the affinity for conventional PKCs rather than for novel PKCs, and the introduction of a methyl group into position 10 would enhance the affinity for both conventional and novel PKCs. Activation of PKCα is suggested to be involved in cancer cell growth, and PKCδ to play a tumor suppressor role and to be involved in apoptosis.[6]
Available forms
Aplog-1, a simple and less hydrophobic analog of aplysiatoxin, is a PKC ligand with little tumor-promoting activity that showed growth-inhibitory activities against several cancer cell lines. Multiple derivatives were evaluated for their antiproliferative activity against several human cancer cell lines and binding activity for PKCδ, which plays a tumor suppressor role and is involved in apoptosis. The results showed that the dimethyl groups at position 6 are indispensable to these activities, but that the hydroxyl group at position 18 is not necessary. Of note, the more hydrophobic 12,12-dimethyl-aplog-1 did not show any tumor-promoting activity in vitro or in vivo. These results suggest that hydrophobicity around the spiroketal moiety of aplog-1 would enhance antiproliferative activities but not tumor-promoting activity. The antiproliferative activities of the analogs to not simply depend on molecular hydrophobicity, and the local hydrophobicity around position 10 plays an important role in enhancing antiproliferative activities. Because the analogues possess the skeleton of tumor-promoting aplysiatoxin and debromoaplysiatoxin, the adverse effects would be most likely tumor promotion.[6]
Toxicity
Debromoaplysiatoxin has activity against P-388 mouse lymphatic leukaemia and was found to cause dermatitis. It has been found to be active at concentrations of 0.005 nmol per ear. The compound was first isolated from the digestive tract of the sea hare Stylocheilus longicauda. Accidental skin contact with sea hare toxin extract led to skin irritations. This phenolic bislactones has been shown to have potent tumor promoting activities. Debromoaplysiatoxin produces erythema, blisters and necrosis.
The dehydrated anhydrotoxins of debromoaplysiatoxin are relatively nontoxic. Examination of the structure–activity relationship (SAR) of this hydrophobic region showed that the absence of the brominated molecule in moieties of Lyngbya toxins reduced malignant transformation and DNA synthesis in cells.[7]
Effects on animals
The response of animals to debromoaplysiatoxin is variable. For example, the response of mice with P-388 lymphocytic leukemia to injections of debromoaplysiatoxin. The result was that the mice had good antileukemia response after treatment with debromoaplysiatoxin. The disadvantage was that these responses were only measured by a dose at which nearly 50 percent of the mice died of toxicity (LD50).[8]
Toxicity of aplysiatoxin and debromoaplysiatoxin from G. coronopifolia against mice is also shown in Table 1. Aplysiatoxin was twice as toxic to mice as debromoaplysiatoxin. The characteristic symptom of these toxins in mice was diarrhea, which usually occurred 30 minutes after injection of toxins. Lethargy (a state of tiredness, weariness, fatigue, or lack of energy), muscular contractions and sometimes hind leg paralysis were observed. Death was observed by 1.2 g of each algae in the mouse toxicity assay.[3]
Furthermore, scientists speculate that debromoaplysiatoxin is accumulated in marine organisms by trophic transfer. In particular, trophic transfer of debromoaplysiatoxin from L. majuscula into Stylocheilus striatus (a sea slug) has been established.[9] This indicates that the compound isn't excreted by the animals causing a higher risk of death related to the amount of food consumed by the animal and its age.
Also, dermal diseases in the manatee population is likely to be caused by exposure to Lyngbya. The algal mat samples collected from manatee dorsum were dominated by Lyngbya spp. Other algae were present in very low quantities and varied in composition. Lyngbya-dominated mats were also collected from the walls of manatee holding tanks. Manatee feces was sampled from the anal opening and debromoaplysiatoxin was identified in multiple samples, proving the exposure of manatees to Lyngbya toxins.[10] It is highly plausible that dermal disease in the manatee population is linked with exposure to Lyngbyatoxins. In contrast to humans who would likely have periodic exposure to Lyngbya while swimming, Lyngbya growing on the dorsa of manatees would cause 24 h per day exposure to the toxin. It may be hypothesized that the continual presence of these toxin-producing cyanobacteria on the skin of these clinically ill animals may be contributing to their dermatologic disease.[10] (5)
In rabbits and hairless mice topical application of debromoaplysiatoxin produced severe cutaneous inflammatory reactions. DAT produces dermatitis on the murine ear at a very low dose. This inflammation response and the mechanism of tumor promotion is likely to be mediated through activation of calcium activated, phospholipid-dependent protein kinase C.[11]
In the aplysiatoxin class, debromoaplysiatoxin, aplysiatoxin, and 19-bromoaplysiatoxin have been found to be tumor promoters in mouse skin. Also rat tracheal cells in culture are sensitive indicators for the presence of the polyacetates, aplysiatoxin and debromoaplysiatoxin. It causes increase in colony formation and is in agreement with a proliferative activation ('triggering') of the basal cell population from the normally quiescent Go state found in intact tracheal epithelium.[12][13] The results also suggest that the polyacetates are good candidates for tumor promoters in vivo.
Related Research Articles
Turtle fibropapillomatosis (FP) is a disease of sea turtles. The condition is characterized by benign but ultimately debilitating epithelial tumours on the surface of biological tissues. FP exists all over the world, but it is most prominent in warmer climates, affecting up to 50–70% of some populations.
Bufotalin is a cardiotoxic bufanolide steroid, cardiac glycoside analogue, secreted by a number of toad species. Bufotalin can be extracted from the skin parotoid glands of several types of toad.
Palytoxin, PTX or PLTX is an intense vasoconstrictor, and is considered to be one of the most poisonous non-protein substances known, second only to maitotoxin in terms of toxicity in mice.
Phorbol is a natural, plant-derived organic compound. It is a member of the tigliane family of diterpenes. Phorbol was first isolated in 1934 as the hydrolysis product of croton oil, which is derived from the seeds of the purging croton, Croton tiglium. The structure of phorbol was determined in 1967. Various esters of phorbol have important biological properties, the most notable of which is the capacity to act as tumor promoters through activation of protein kinase C. They mimic diacylglycerols, glycerol derivatives in which two hydroxyl groups have reacted with fatty acids to form esters. The most common and potent phorbol ester is 12-O-tetradecanoylphorbol-13-acetate (TPA), also called phorbol-12-myristate-13-acetate (PMA), which is used as a biomedical research tool in contexts such as models of carcinogenesis.
Pederin is a vesicant toxic amide with two tetrahydropyran rings, found in the haemolymph of the beetle genus Paederus, including the Nairobi fly, belonging to the family Staphylinidae. It was first characterized by processing 25 million field-collected P. fuscipes. It makes up approximately 0.025% of an insects weight.
Halomon is a polyhalogenated monoterpene first isolated from the marine red algae Portieria hornemannii. Halomon has attracted research interest because of its promising profile of selective cytotoxicity that suggests its potential use as an antitumor agent.
The PHLPP isoforms are a pair of protein phosphatases, PHLPP1 and PHLPP2, that are important regulators of Akt serine-threonine kinases and conventional/novel protein kinase C (PKC) isoforms. PHLPP may act as a tumor suppressor in several types of cancer due to its ability to block growth factor-induced signaling in cancer cells.
Cylindrospermopsin is a cyanotoxin produced by a variety of freshwater cyanobacteria. CYN is a polycyclic uracil derivative containing guanidino and sulfate groups. It is also zwitterionic, making it highly water soluble. CYN is toxic to liver and kidney tissue and is thought to inhibit protein synthesis and to covalently modify DNA and/or RNA. It is not known whether cylindrospermopsin is a carcinogen, but it appears to have no tumour initiating activity in mice.
Aplysiatoxin is a cyanotoxin produced by certain cyanobacteria species. It is used as a defensive secretion to protect these cyanobacteria from predation by fish, being a potent irritant and carcinogen, by acting as a powerful activator of protein kinase C. While this action has a tumour-promoting effect, protein kinase C activation can be medically beneficial for some other applications, and synthetic analogues of aplysiatoxin have been researched for anti-cancer effects.
Nodularins are potent toxins produced by the cyanobacterium Nodularia spumigena, among others. This aquatic, photosynthetic cyanobacterium forms visible colonies that present as algal blooms in brackish water bodies throughout the world. The late summer blooms of Nodularia spumigena are among the largest cyanobacterial mass occurrences in the world. Cyanobacteria are composed of many toxic substances, most notably of microcystins and nodularins: the two are not easily differentiated. A significant homology of structure and function exists between the two, and microcystins have been studied in greater detail. Because of this, facts from microcystins are often extended to nodularins.
Dogger Bank itch is a cutaneous condition characterized by a long-lasting dermatitis caused by exposure to the sea chervil, Alcyonidium diaphanum, a bryozoan. The disease, common in fishermen who work in the North Sea, has been recognized by the Danish Workman's Compensation Act since 1939.
Microcystin-LR (MC-LR) is a toxin produced by cyanobacteria. It is the most toxic of the microcystins.
Lyngbyatoxin-a is a cyanotoxin produced by certain cyanobacteria species, most notably Moorea producens. It is produced as defense mechanism to ward off any would-be predators of the bacterium, being a potent blister agent as well as carcinogen. Low concentrations cause a common skin condition known as seaweed dermatitis.
Mezerein is a toxic diterpene ester found in the sap of Daphne mezereum and related plants. Plants of the genera Euphorbiaceae and Thymelaeaceae possess a wide variety of different phorbol esters, which share the capacity of mimicking diacylglycerol (DAG) and thus activating different isoforms of protein kinase C. Mezerein was first isolated in 1975. It has antileukemic properties in mice, but it is also defined as a weak promoter of skin cancers in the same species. All parts of the plants contain an acrid and irritant sap that contains mezerein, thought to be the principal poison. The sap is especially prevalent in the bark and berries.
Curacin A is a hybrid polyketide synthase (PKS)/nonribosomal peptide synthase (NRPS) derived natural product produced isolated from the cyanobacterium Lyngbya majuscula. Curacin A belongs to a family of natural products including jamaicamide, mupirocin, and pederin that have an unusual terminal alkene. Additionally, Curacin A contains a notable thiazoline ring and a unique cyclopropyl moiety, which is essential to the compound's biological activity. Curacin A has been characterized as potent antiproliferative cytotoxic compound with notable anticancer activity for several cancer lines including renal, colon, and breast cancer. Curacin A has been shown to interact with colchicine binding sites on tubulin, which inhibits microtubule polymerization, an essential process for cell division and proliferation.
Antillatoxin (ATX) is a potent lipopeptide neurotoxin produced by the marine cyanobacterium Lyngbya majuscula. ATX activates voltage-gated sodium channels, which can cause cell depolarisation, NMDA-receptor overactivity, excess calcium influx and neuronal necrosis.
Kalkitoxin, a toxin derived from the cyanobacterium Lyngbya majuscula, induces NMDA receptor mediated neuronal necrosis, blocks voltage-dependent sodium channels, and induces cellular hypoxia by inhibiting the electron transport chain (ETC) complex 1.
Crotoxin (CTX) is the main toxic compound in the snake venom of the South American rattlesnake, Crotalus durissus terrificus. Crotoxin is a heterodimeric beta-neurotoxin, composed of an acidic, non-toxic and non-enzymatic subunit (CA), and a basic, weakly toxic, phospholipase A2 protein (CB). This neurotoxin causes paralysis by both pre- and postsynaptic blocking of acetylcholine signalling.
Jamaicamide A is a lipopeptide isolated from the cyanobacterium Moorea producens, formerly known as Lyngbya majuscula. Jamaicamide A belongs to a family of compounds collectively called jamaicamides, which are sodium channel blockers with potent neurotoxicity in a cellular model. Jamaicamide A has several unusual functionalities, including an alkynyl bromide, vinyl chloride, β-methoxy eneone system, and pyrrolinone ring.
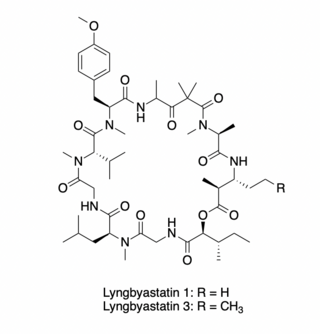
Lyngbyastatins 1 and 3 are cytotoxic cyclic depsipeptides that possess antiproliferative activity against human cancer cell lines. These compounds, first isolated from the extract of a Lyngbya majuscula/Schizothrix calcicola assemblage and from L. majuscula Harvey ex Gomont (Oscillatoriaceae) strains, respectively, target the actin cytoskeleton of eukaryotic cells.
References
↑ Hashimoto, Y.; Kamiya, H.; Yamazato, K.; Nozawa (1975). "Occurrence of a toxic blue-green alga inducing skin dermatitis in okinawa". Toxicon. 13 (2): 95–96. doi:10.1016/0041-0101(75)90034-3.
1 2 Fujiki, H.; Ikegami, K.; Hakii, H.; Suganuma, M.; Yamaizumi, Z.; Yamazato, K.; Moore, R.E.; Sugimara, T. (1985). "A blue-green alga from Okinawa contains aplysiatoxins, the third class of tumor promoters". Japanese Journal of Cancer Research. 76 (4): 257–259.
1 2 Nagai, H.; Yasumoto, T.; Hokama, Y. (July 1996). "Aplysiatoxin and debromoaplysiatoxin as the causative agents of a red alga Gracilaria coronopifolia poisoning in Hawaii". Toxicon. 34 (7): 753–761. doi:10.1016/0041-0101(96)00014-1.
↑ Okamura, H.; Kuroda, S.; Ikegami, S.; Tomita, K.; Sugimoto, Y.; Sakaguchi, S.; Ito, Y.; Katsuki, T.; Yamaguchi, M. (November 1993). "A Formal Synthesis of Aplysiatoxin - Enantioselective Synthesis of Kishi Aldehyde". Tetrahedron. 49 (46): 10531–10554. doi:10.1016/s0040-4020(01)81547-7.
1 2 Apsimon, John (February 1992). The Total Synthesis of Natural Products. Wiley-Interscience.
1 2 Kikumori, M.; Yanagita, R.C.; Tokuda, H.; Suzuki, N.; Nagai, H.; Suenaga, K.; Irie, K. (June 2012). "Structure–Activity Studies on the Spiroketal Moiety of a Simplified Analogue of Debromoaplysiatoxin with Antiproliferative Activity". Journal of Medicinal Chemistry. 55 (11): 5614–5626. doi:10.1021/jm300566h. PMID22625994.
↑ Osborne, N.J.T.; Webb, P.M.; Shaw, G.R. (November 2001). "The toxins of Lyngbya majuscula and their human and ecological health effects". Environment International. 27 (5): 381–392. doi:10.1016/s0160-4120(01)00098-8. PMID11757852.
↑ Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. (1977). "Antileukemia Activity in the Oscillatoriaceae: Isolation of Debromoaplysiatoxin from Lyngbya". Science. 196 (4289): 538–540. doi:10.1126/science.403608.
↑ Capper, A.; Tibbetts, I.R.; O'Neil, J.M.; Shaw, G.R. (July 2005). "The fate of Lyngbya majuscula toxins in three potential consumers". Journal of Chemical Ecology. 31 (7): 1595–1606. doi:10.1007/s10886-005-5800-5.
1 2 Harr, K.E.; Szabo, N.J.; Cichra, M.; Phlips, E.J. (August 2008). "Debromoaplysiatoxin in Lyngbya-dominated mats on manatees (Trichechus manatus latirostris) in the Florida King's Bay ecosystem". Toxicon. 52 (2): 385–388. doi:10.1016/j.toxicon.2008.05.016. PMID18585400.
↑ Solomon, A.E.; Stoughton, R.B. (1978). "Dermatitis from purified sea algae toxin (debromoaplysiatoxin)". Archives of Dermatology. 114 (9): 1333–1335. doi:10.1001/archderm.114.9.1333.
↑ Horowitz, A.D.; Fujiki, H.; Weinstein, I.B.; Jeffrey, A.; Okin, E.; Moore, R.E.; Sugimara, T. (1983). "Comparative Effects of Aplysiatoxin, Debromoaplysiatoxin, and Teleocidin on Receptor Binding and Phospholipid Metabolism". Cancer Research. 43 (4): 1529–1535.
↑ Mass, M.J.; Lasley, J.A.; Marr, C.M.; Arnold, J.T.; Steele, V.E. (January 1987). "Colony formation enhancement of rat tracheal and nasal epithelial cells by polyacetate, indole alkaloid, and phorbol ester tumor promoters". Carcinogenesis. 8 (1): 179–181. doi:10.1093/carcin/8.1.179.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.