
Holoprosencephaly (HPE) is a cephalic disorder in which the prosencephalon fails to develop into two hemispheres, typically occurring between the 18th and 28th day of gestation. Normally, the forebrain is formed and the face begins to develop in the fifth and sixth weeks of human pregnancy. The condition also occurs in other species.
Alien hand syndrome (AHS) or Dr. Strangelove syndrome is a category of conditions in which a person experiences their limbs acting seemingly on their own, without conscious control over the actions. There are a variety of clinical conditions that fall under this category, most commonly affecting the left hand. There are many similar terms for the various forms of the condition, but they are often used inappropriately. The affected person may sometimes reach for objects and manipulate them without wanting to do so, even to the point of having to use the controllable hand to restrain the alien hand. The occurrence of alien hand syndrome can be usefully conceptualized as a phenomenon reflecting a functional "disentanglement" between thought and action.
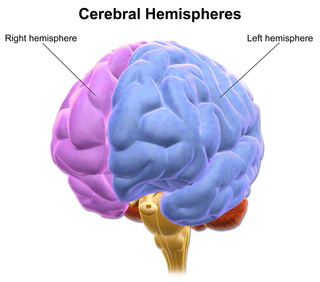
The vertebrate cerebrum (brain) is formed by two cerebral hemispheres that are separated by a groove, the longitudinal fissure. The brain can thus be described as being divided into left and right cerebral hemispheres. Each of these hemispheres has an outer layer of grey matter, the cerebral cortex, that is supported by an inner layer of white matter. In eutherian (placental) mammals, the hemispheres are linked by the corpus callosum, a very large bundle of nerve fibers. Smaller commissures, including the anterior commissure, the posterior commissure and the fornix, also join the hemispheres and these are also present in other vertebrates. These commissures transfer information between the two hemispheres to coordinate localized functions.
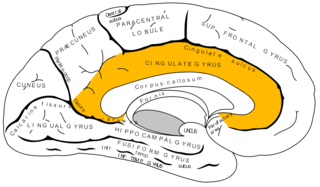
The cingulate cortex is a part of the brain situated in the medial aspect of the cerebral cortex. The cingulate cortex includes the entire cingulate gyrus, which lies immediately above the corpus callosum, and the continuation of this in the cingulate sulcus. The cingulate cortex is usually considered part of the limbic lobe.
Split-brain or callosal syndrome is a type of disconnection syndrome when the corpus callosum connecting the two hemispheres of the brain is severed to some degree. It is an association of symptoms produced by disruption of, or interference with, the connection between the hemispheres of the brain. The surgical operation to produce this condition involves transection of the corpus callosum, and is usually a last resort to treat refractory epilepsy. Initially, partial callosotomies are performed; if this operation does not succeed, a complete callosotomy is performed to mitigate the risk of accidental physical injury by reducing the severity and violence of epileptic seizures. Before using callosotomies, epilepsy is instead treated through pharmaceutical means. After surgery, neuropsychological assessments are often performed.

The longitudinal fissure is the deep groove that separates the two cerebral hemispheres of the vertebrate brain. Lying within it is a continuation of the dura mater called the falx cerebri. The inner surfaces of the two hemispheres are convoluted by gyri and sulci just as is the outer surface of the brain.

Diffuse axonal injury (DAI) is a brain injury in which scattered lesions occur over a widespread area in white matter tracts as well as grey matter. DAI is one of the most common and devastating types of traumatic brain injury and is a major cause of unconsciousness and persistent vegetative state after severe head trauma. It occurs in about half of all cases of severe head trauma and may be the primary damage that occurs in concussion. The outcome is frequently coma, with over 90% of patients with severe DAI never regaining consciousness. Those who awaken from the coma often remain significantly impaired.
Neuroscience and intelligence refers to the various neurological factors that are partly responsible for the variation of intelligence within species or between different species. A large amount of research in this area has been focused on the neural basis of human intelligence. Historic approaches to studying the neuroscience of intelligence consisted of correlating external head parameters, for example head circumference, to intelligence. Post-mortem measures of brain weight and brain volume have also been used. More recent methodologies focus on examining correlates of intelligence within the living brain using techniques such as magnetic resonance imaging (MRI), functional MRI (fMRI), electroencephalography (EEG), positron emission tomography and other non-invasive measures of brain structure and activity.

The angular gyrus is a region of the brain lying mainly in the posteroinferior region of the parietal lobe, occupying the posterior part of the inferior parietal lobule. It represents the Brodmann area 39.
Cerebral atrophy is a common feature of many of the diseases that affect the brain. Atrophy of any tissue means a decrement in the size of the cell, which can be due to progressive loss of cytoplasmic proteins. In brain tissue, atrophy describes a loss of neurons and the connections between them. Brain atrophy can be classified into two main categories: generalized and focal atrophy. Generalized atrophy occurs across the entire brain whereas focal atrophy affects cells in a specific location. If the cerebral hemispheres are affected, conscious thought and voluntary processes may be impaired.

The anterior cerebral artery (ACA) is one of a pair of cerebral arteries that supplies oxygenated blood to most midline portions of the frontal lobes and superior medial parietal lobes of the brain. The two anterior cerebral arteries arise from the internal carotid artery and are part of the circle of Willis. The left and right anterior cerebral arteries are connected by the anterior communicating artery.

A corpus callosotomy is a palliative surgical procedure for the treatment of medically refractory epilepsy. The procedure was first performed in 1940 by William P. van Wagenen. In this procedure, the corpus callosum is cut through, in an effort to limit the spread of epileptic activity between the two halves of the brain. Another method to treat epilepsy is vagus nerve stimulation.
Agenesis of the corpus callosum (ACC) is a rare birth defect in which there is a complete or partial absence of the corpus callosum. It occurs when the development of the corpus callosum, the band of white matter connecting the two hemispheres in the brain, in the embryo is disrupted. The result of this is that the fibers that would otherwise form the corpus callosum are instead longitudinally oriented along the ipsilateral ventricular wall and form structures called Probst bundles.
The posterior cerebral artery (PCA) is one of a pair of cerebral arteries that supply oxygenated blood to the occipital lobe, part of the back of the human brain. The two arteries originate from the distal end of the basilar artery, where it bifurcates into the left and right posterior cerebral arteries. These anastomose with the middle cerebral arteries and internal carotid arteries via the posterior communicating arteries.
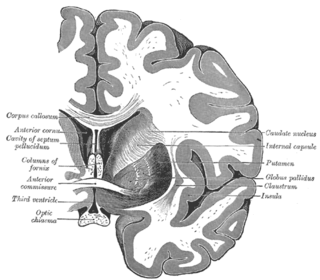
The anterior commissure is a white matter tract connecting the two temporal lobes of the cerebral hemispheres across the midline, and placed in front of the columns of the fornix. In all but five species of mammal the great majority of fibers connecting the two hemispheres travel through the corpus callosum, which in humans and all non-monotremes is more than 10 times larger than the anterior commissure. Other routes of communication pass through the hippocampal commissure or, indirectly, via subcortical connections. Nevertheless, the anterior commissure is a significant pathway that can be clearly distinguished in the brains of all mammals.
The commissural fibers or transverse fibers are axons that connect the two hemispheres of the brain. Huge numbers of commissural fibers make up the commissural tracts in the brain, the largest of which is the corpus callosum.
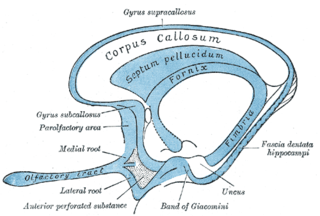
The indusium griseum, consists of a thin membranous layer of grey matter in contact with the upper surface of the corpus callosum and continuous laterally with the grey matter of the cingulate cortex and inferiorly with the hippocampus. It is vestigial in humans and is a remnant of the former position of the hippocampus in lower animals.
Longitudinal callosal fascicles, or Probst bundles, are aberrant bundles of axons that run in a front-back (antero-posterior) direction rather than a left-right direction between the cerebral hemispheres. They are characteristic of patients with agenesis of the corpus callosum and are due to failure of the callosally-projecting neurons to extend axons across the midline and therefore form the corpus callosum. The inability of these axons to cross the midline results in anomalous axonal guidance and front-to-back projections within each hemisphere, rather than connecting between the hemispheres in the normal corpus callosum.

Disconnection syndrome is a general term for a collection of neurological symptoms caused – via lesions to associational or commissural nerve fibres – by damage to the white matter axons of communication pathways in the cerebrum, independent of any lesions to the cortex. The behavioral effects of such disconnections are relatively predictable in adults. Disconnection syndromes usually reflect circumstances where regions A and B still have their functional specializations except in domains that depend on the interconnections between the two regions.

Francisco Aboitiz is a Chilean neuroscientist, academic, and author. He is a professor at the Medical School and the Director of the Interdisciplinary Center for Neuroscience NeuroUC at Pontificia Universidad Católica (PUC) de Chile.









