Bacteriorhodopsin (Bop) is a protein used by Archaea, most notably by haloarchaea, a class of the Euryarchaeota.[1] It acts as a proton pump; that is, it captures light energy and uses it to move protons across the membrane out of the cell.[2] The resulting proton gradient is subsequently converted into chemical energy.[3]
Bacteriorhodopsin is a light-driven H+ ion transporter found in some haloarchaea, most notably Halobacterium salinarum (formerly known as syn. H. halobium). The proton-motive force generated by the protein is used by ATP synthase to generate adenosine triphosphate (ATP). By expressing bacteriorhodopsin, the archaea cells are able to synthesise ATP in the absence of a carbon source.[4][5]
Structure
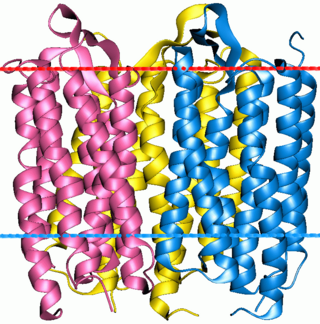
A bacteriorhodopsin trimer, showing the approximate positions of the extracellular and cytoplasmic sides of the membrane (red and blue lines respectively)
Bacteriorhodopsin is a 27 kDaintegral membrane protein usually found in two-dimensional crystalline patches known as "purple membrane", which can occupy almost 50% of the surface area of the archaeal cell. The repeating element of the hexagonal lattice is composed of three identical protein chains, each rotated by 120 degrees relative to the others.[6] Each monomer has seven transmembrane alpha helices and an extracellular-facing, two-stranded beta sheet.[7][8]
Bacteriorhodopsin molecule is purple and is most efficient at absorbing green light (in the wavelength range 500-650 nm). In the native membrane, the protein has a maximum absorbance at 553 nm, however addition of detergent disrupts the trimeric form, leading a loss of exciton coupling between the chromophores, and the monomeric form consequently has an absorption maximum of 568 nm.[13][14]
Bacteriorhodopsin has a broad excitation spectrum. For a detection wavelength between 700 and 800nm, it has an appreciable detected emission for excitation wavelengths between 470nm and 650nm (with a peak at 570nm).[15] When pumped at 633nm, the emission spectrum has appreciable intensity between 650nm and 850nm.[16]
Mechanism
Photocycle overview
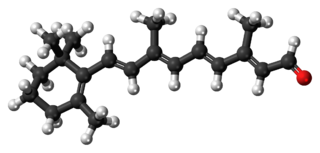
Bacteriorhodopsin is a light-driven proton pump. It is the retinal molecule that changes its isomerization state from all-trans to 13-cis when it absorbs a photon. The surrounding protein responds to the change in the chromophore shape, by undergoing an ordered sequence of conformational changes (collectively known as the photocycle).[17] The conformational changes alter the pKa values of conserved amino acids in the core of the protein, including Asp85, Asp96 and the Schiff base N atom (Lys216). These sequential changes in acid dissociation constant, result in the transfer of one proton from the intracellular side to the extracellular side of the membrane for each photon absorbed by the chromophore.
The bacteriorhodopsin photocycle consists of nine distinct stages, starting from the ground or resting state, which is denoted 'bR'. The intermediates are identified by single letters and may be distinguished by their absorption spectra.[18] The nine stages are:
bR + photon → K ⇌ L ⇌ M1⇌ M2⇌ M2' ⇌ N ⇌ N' ⇌ O ⇌ bR[18]
Ground state + photon → K state → L state
Conformational change, paired stereogram. The orange molecule is all-trans retinal and the red molecule is 13-cis retinal.
Bacteriorhodopsin in the ground state absorbs a photon and the retinal changes isomerization from all-trans 15-anti to the strained 13-cis 15-anti in the K state. The isomerisation reaction is fast and occurs in less than 1 ps. The retinal adopts a less strained conformation to form the L intermediate.
L state → M1 state
Asp85 accepts a proton from the Schiff base N atom. In the M1 intermediate, neither the Schiff base nor Asp85 are charged.
M1 state → M2 state
The Schiff base rotates away from the extracellular side of the protein towards the cytoplasmic side, in preparation to accept a new proton.
M2 state → M2' state
A proton is released from Glu204 and Glu194 to the extracellular medium.
M2' state → N state
The retinal Schiff base accepts a proton from Asp96. In the N state, both Asp96 and the Schiff base are charged.
N state → N' state
Asp96 accepts a proton from the cytoplasmic side of the membrane and becomes uncharged.
N' state → O state
Retinal reisomerizes to the all-trans state.
O state → ground state
Asp85 transfers a proton to Glu194 and Glu204 [19][20] on the extracellular face of the protein.
Bacteriorhodopsin is similar to vertebraterhodopsins, the pigments that sense light in the retina. Rhodopsins also contain retinal; however, the functions of rhodopsin and bacteriorhodopsin are different, and there is limited similarity in their amino acid sequences. Both rhodopsin and bacteriorhodopsin belong to the 7TM receptor family of proteins, but rhodopsin is a G protein-coupled receptor and bacteriorhodopsin is not. In the first use of electron crystallography to obtain an atomic-level protein structure, the structure of bacteriorhodopsin was resolved in 1990.[22] It was then used as a template to build models of G protein-coupled receptors before crystallographic structures were also available for these proteins. It has been excessively studied on both mica[23][24] and glass substrates using Atomic force microscopy and Femtosecond crystallography.[25]
All other phototrophic systems in bacteria, algae, and plants use chlorophylls or bacteriochlorophylls rather than bacteriorhodopsin. These also produce a proton gradient, but in a quite different and more indirect way involving an electron transfer chain consisting of several other proteins. Furthermore, chlorophylls are aided in capturing light energy by other pigments known as "antennas"; these are not present in bacteriorhodopsin-based systems. It is possible that phototrophy independently evolved at least twice, once in bacteria and once in archaea.
↑ Pebay-Peroua E, Rummel G, Rosenbusch JP, Landau EM (1997). "X-ray structure of bacteriorhodopsin at 2.5 Å from microcrystals grown in lipidic cubic phases". Science. 277 (5332): 1676–1681. doi:10.1126/science.277.5332.1676. PMID9287223.
↑ Luecke H, Schobert B, Richter HT, Cartailler JP, Lanyi JK (1999). "Structure of bacteriorhodopsin at 1.55 Å resolution". Journal of Molecular Biology. 291 (4): 899–911. doi:10.1006/jmbi.1999.3027. PMID10452895.
↑ Oesterhelt, Dieter (1982). "[3] Reconstitution of the retinal proteins bacteriorhodopsin and halorhodopsin". Reconstitution of the retinal proteins bacteriorhodopsin and halorhodopsin. Methods in Enzymology. Vol.88. pp.10–17. doi:10.1016/0076-6879(82)88006-3. ISBN9780121819880.
↑ Pescitelli G, Woody RW (2012). "The Exciton Origin of the Visible Circular Dichroism Spectrum of Bacteriorhodopsin". Journal of Physical Chemistry B. 116 (23): 6751–6763. doi:10.1021/jp212166k. PMID22329810.
↑ Schenkl, Selma; Zgrablic, Goran; Portuondo-Campa, Erwin; Haacke, Stefan; Chergui, Majed (2007). "On the excitation wavelength dependence of the fluorescence of bacteriorhodopsin". Chemical Physics Letters. 441 (4–6): 322–326. Bibcode:2007CPL...441..322S. doi:10.1016/j.cplett.2007.04.086.
↑ Ohtani, H.; Tsukamoto, Y.; Sakoda, Y.; Hamaguchi, H. (1995). "Fluorescence spectra of bacteriorhodopsin and the intermediates O and Q at room temperature". FEBS Lett. 359 (1): 65–68. doi:10.1016/0014-5793(94)01440-c. PMID7851532.
↑ Henderson R, Baldwin JM, Ceska TA, Zemlin F, Beckmann E, Downing KH (1990). "Model for the structure of bacteriorhodopsin based on high-resolution electron cryo-microscopy". J Mol Biol. 213 (4): 899–929. doi:10.1016/S0022-2836(05)80271-2. PMID2359127.{{cite journal}}: CS1 maint: multiple names: authors list (link)
1 2 Nishikawa, T.; Murakami, M. (2005). "Crystal structure of the 13-cis isomer of bacteriorhodopsin in the dark-adapted state". J. Mol. Biol. 352 (2): 319–328. doi:10.1016/j.jmb.2005.07.021. PMID16084526. PDB ID: 1X0S.
1 2 Image created with RasTop (Molecular Visualization Software).
The green fluorescent protein (GFP) is a protein that exhibits bright green fluorescence when exposed to light in the blue to ultraviolet range. The label GFP traditionally refers to the protein first isolated from the jellyfish Aequorea victoria and is sometimes called avGFP. However, GFPs have been found in other organisms including corals, sea anemones, zoanithids, copepods and lancelets.
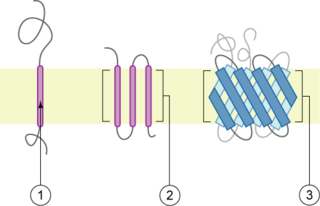
A transmembrane protein (TP) is a type of integral membrane protein that spans the entirety of the cell membrane. Many transmembrane proteins function as gateways to permit the transport of specific substances across the membrane. They frequently undergo significant conformational changes to move a substance through the membrane. They are usually highly hydrophobic and aggregate and precipitate in water. They require detergents or nonpolar solvents for extraction, although some of them (beta-barrels) can be also extracted using denaturing agents.
Rhodopsin, also known as visual purple, is a protein encoded by the RHO gene and a G-protein-coupled receptor (GPCR). It is the opsin of the rod cells in the retina and a light-sensitive receptor protein that triggers visual phototransduction in rods. Rhodopsin mediates dim light vision and thus is extremely sensitive to light. When rhodopsin is exposed to light, it immediately photobleaches. In humans, it is regenerated fully in about 30 minutes, after which the rods are more sensitive. Defects in the rhodopsin gene cause eye diseases such as retinitis pigmentosa and congenital stationary night blindness.
Retinal is a polyene chromophore. Retinal, bound to proteins called opsins, is the chemical basis of visual phototransduction, the light-detection stage of visual perception (vision).
Proteorhodopsin is a family of transmembrane proteins that use retinal as a chromophore for light-mediated functionality, in this case, a proton pump. pRhodopsin is found in marine planktonic bacteria, archaea and eukaryotes (protae), but was first discovered in bacteria.
Animal opsins are G-protein-coupled receptors and a group of proteins made light-sensitive via a chromophore, typically retinal. When bound to retinal, opsins become Retinylidene proteins, but are usually still called opsins regardless. Most prominently, they are found in photoreceptor cells of the retina. Five classical groups of opsins are involved in vision, mediating the conversion of a photon of light into an electrochemical signal, the first step in the visual transduction cascade. Another opsin found in the mammalian retina, melanopsin, is involved in circadian rhythms and pupillary reflex but not in vision. Humans have in total nine opsins. Beside vision and light perception, opsins may also sense temperature, sound, or chemicals.
Channelrhodopsins are a subfamily of retinylidene proteins (rhodopsins) that function as light-gated ion channels. They serve as sensory photoreceptors in unicellular green algae, controlling phototaxis: movement in response to light. Expressed in cells of other organisms, they enable light to control electrical excitability, intracellular acidity, calcium influx, and other cellular processes. Channelrhodopsin-1 (ChR1) and Channelrhodopsin-2 (ChR2) from the model organism Chlamydomonas reinhardtii are the first discovered channelrhodopsins. Variants that are sensitive to different colors of light or selective for specific ions have been cloned from other species of algae and protists.
An electrochemical gradient is a gradient of electrochemical potential, usually for an ion that can move across a membrane. The gradient consists of two parts:
Halorhodopsin is a seven-transmembrane retinylidene protein from microbial rhodopsin family. It is a chloride-specific light-gated ion pump found in archaea known as halobacteria. It is activated by green light wavelengths of approximately 578nm. Halorhodopsin also shares sequence similarity to channelrhodopsin, another light-driven ion channel.
Photoreceptor proteins are light-sensitive proteins involved in the sensing and response to light in a variety of organisms. Some examples are rhodopsin in the photoreceptor cells of the vertebrate retina, phytochrome in plants, and bacteriorhodopsin and bacteriophytochromes in some bacteria. They mediate light responses as varied as visual perception, phototropism and phototaxis, as well as responses to light-dark cycles such as circadian rhythm and other photoperiodisms including control of flowering times in plants and mating seasons in animals.
Retinylidene proteins, or rhodopsins in a broad sense, are proteins that use retinal as a chromophore for light reception. They are the molecular basis for a variety of light-sensing systems from phototaxis in flagellates to eyesight in animals. Retinylidene proteins include all forms of opsin and rhodopsin. While rhodopsin in the narrow sense refers to a dim-light visual pigment found in vertebrates, usually on rod cells, rhodopsin in the broad sense refers to any molecule consisting of an opsin and a retinal chromophore in the ground state. When activated by light, the chromophore is isomerized, at which point the molecule as a whole is no longer rhodopsin, but a related molecule such as metarhodopsin. However, it remains a retinylidene protein. The chromophore then separates from the opsin, at which point the bare opsin is a retinylidene protein. Thus, the molecule remains a retinylidene protein throughout the phototransduction cycle.
A retinalophototroph is one of two different types of phototrophs, and are named for retinal-binding proteins they utilize for cell signaling and converting light into energy. Like all photoautotrophs, retinalophototrophs absorb photons to initiate their cellular processes. However, unlike all photoautotrophs, retinalophototrophs do not use chlorophyll or an electron transport chain to power their chemical reactions. This means retinalophototrophs are incapable of traditional carbon fixation, a fundamental photosynthetic process that transforms inorganic carbon into organic compounds. For this reason, experts consider them to be less efficient than their chlorophyll-using counterparts, chlorophototrophs.
The visual cycle is a process in the retina that replenishes the molecule retinal for its use in vision. Retinal is the chromophore of most visual opsins, meaning it captures the photons to begin the phototransduction cascade. When the photon is absorbed, the 11-cis retinal photoisomerizes into all-trans retinal as it is ejected from the opsin protein. Each molecule of retinal must travel from the photoreceptor cell to the RPE and back in order to be refreshed and combined with another opsin. This closed enzymatic pathway of 11-cis retinal is sometimes called Wald's visual cycle after George Wald (1906–1997), who received the Nobel Prize in 1967 for his work towards its discovery.
Peropsin, a visual pigment-like receptor, is a protein that in humans is encoded by the RRH gene. It belongs like other animal opsins to the G protein-coupled receptors. Even so, the first peropsins were already discovered in mice and humans in 1997, not much is known about them.
Peter Hegemann is a Hertie Senior Research Chair for Neurosciences and a Professor of Experimental Biophysics at the Department of Biology, Faculty of Life Sciences, Humboldt University of Berlin, Germany. He is known for his discovery of channelrhodopsin, a type of ion channels regulated by light, thereby serving as a light sensor. This created the field of optogenetics, a technique that controls the activities of specific neurons by applying light. He has received numerous accolades, including the Rumford Prize, the Shaw Prize in Life Science and Medicine, and the Albert Lasker Award for Basic Medical Research.
Microbial rhodopsins, also known as bacterial rhodopsins, are retinal-binding proteins that provide light-dependent ion transport and sensory functions in halophilic and other bacteria. They are integral membrane proteins with seven transmembrane helices, the last of which contains the attachment point for retinal.
The lysosomal cystine transporter (LCT) family is part of the TOG Superfamily and includes secondary transport proteins that are derived from animals, plants, fungi and other eukaryotes. They exhibit 7 putative transmembrane α-helical spanners (TMSs) and vary in size between about 200 and 500 amino acyl residues, although most have between 300 and 400 residues.
Archaerhodopsin proteins are a family of retinal-containing photoreceptors found in the archaea genera Halobacterium and Halorubrum. Like the homologous bacteriorhodopsin (bR) protein, archaerhodopsins harvest energy from sunlight to pump H+ ions out of the cell, establishing a proton motive force that is used for ATP synthesis. They have some structural similarities to the mammalian GPCR protein rhodopsin, but are not true homologs.
In molecular biology, the PYP domain is a p-coumaric acid-binding protein domain. They are present in various proteins in bacteria.
Dieter Oesterhelt was a German biochemist. From 1980 until 2008, he was director of the Max Planck Institute for Biochemistry, Martinsried.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.