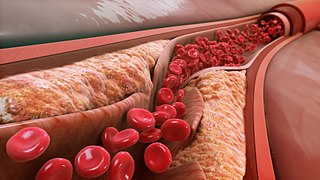
Low-density lipoprotein (LDL) is one of the five major groups of lipoprotein that transport all fat molecules around the body in extracellular water. These groups, from least dense to most dense, are chylomicrons, very low-density lipoprotein (VLDL), intermediate-density lipoprotein (IDL), low-density lipoprotein (LDL) and high-density lipoprotein (HDL). LDL delivers fat molecules to cells. LDL has been associated with the progression of atherosclerosis.
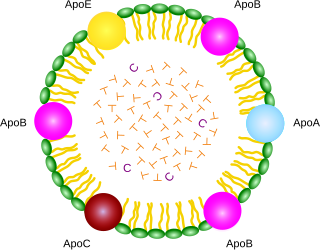
A lipoprotein is a biochemical assembly whose primary function is to transport hydrophobic lipid molecules in water, as in blood plasma or other extracellular fluids. They consist of a triglyceride and cholesterol center, surrounded by a phospholipid outer shell, with the hydrophilic portions oriented outward toward the surrounding water and lipophilic portions oriented inward toward the lipid center. A special kind of protein, called apolipoprotein, is embedded in the outer shell, both stabilising the complex and giving it a functional identity that determines its role.
Very-low-density lipoprotein (VLDL), density relative to extracellular water, is a type of lipoprotein made by the liver. VLDL is one of the five major groups of lipoproteins that enable fats and cholesterol to move within the water-based solution of the bloodstream. VLDL is assembled in the liver from triglycerides, cholesterol, and apolipoproteins. VLDL is converted in the bloodstream to low-density lipoprotein (LDL) and intermediate-density lipoprotein (IDL). VLDL particles have a diameter of 30–80 nanometers (nm). VLDL transports endogenous products, whereas chylomicrons transport exogenous (dietary) products. In the early 2010s both the lipid composition and protein composition of this lipoprotein were characterised in great detail.
Apolipoproteins are proteins that bind lipids to form lipoproteins. They transport lipids in blood, cerebrospinal fluid and lymph.
Hyperlipidemia is abnormally high levels of any or all lipids or lipoproteins in the blood. The term hyperlipidemia refers to the laboratory finding itself and is also used as an umbrella term covering any of various acquired or genetic disorders that result in that finding. Hyperlipidemia represents a subset of dyslipidemia and a superset of hypercholesterolemia. Hyperlipidemia is usually chronic and requires ongoing medication to control blood lipid levels.

Cholesteryl ester transfer protein (CETP), also called plasma lipid transfer protein, is a plasma protein that facilitates the transport of cholesteryl esters and triglycerides between the lipoproteins. It collects triglycerides from very-low-density (VLDL) or Chylomicrons and exchanges them for cholesteryl esters from high-density lipoproteins (HDL), and vice versa. Most of the time, however, CETP does a heteroexchange, trading a triglyceride for a cholesteryl ester or a cholesteryl ester for a triglyceride.

Apolipoprotein E (Apo-E) is a protein involved in the metabolism of fats in the body of mammals. A subtype is implicated in Alzheimer's disease and cardiovascular diseases. It is encoded in humans by the gene APOE.

Apolipoprotein B (ApoB) is a protein that in humans is encoded by the APOB gene. It is commonly used to detect risk of atherosclerotic cardiovascular disease.
Lecithin cholesterol acyltransferase deficiency is a disorder of lipoprotein metabolism. The disease has two forms: Familial LCAT deficiency, in which there is complete LCAT deficiency, and Fish-eye disease, in which there is a partial deficiency.

Apolipoprotein AI(Apo-AI) is a protein that in humans is encoded by the APOA1 gene. As the major component of HDL particles, it has a specific role in lipid metabolism.
Apolipoprotein C-III also known as apo-CIII, and apolipoprotein C3, is a protein that in humans is encoded by the APOC3 gene. Apo-CIII is secreted by the liver as well as the small intestine, and is found on triglyceride-rich lipoproteins such as chylomicrons, very low density lipoprotein (VLDL), and remnant cholesterol.

Apolipoprotein C-IV, also known as apolipoprotein C4, is a protein that in humans is encoded by the APOC4 gene.
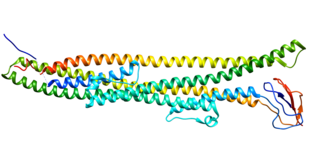
Hepatic lipase (HL), also called hepatic triglyceride lipase (HTGL) or LIPC (for "lipase, hepatic"), is a form of lipase, catalyzing the hydrolysis of triacylglyceride. Hepatic lipase is coded by chromosome 15 and its gene is also often referred to as HTGL or LIPC. Hepatic lipase is expressed mainly in liver cells, known as hepatocytes, and endothelial cells of the liver. The hepatic lipase can either remain attached to the liver or can unbind from the liver endothelial cells and is free to enter the body's circulation system. When bound on the endothelial cells of the liver, it is often found bound to heparan sulfate proteoglycans (HSPG), keeping HL inactive and unable to bind to HDL (high-density lipoprotein) or IDL (intermediate-density lipoprotein). When it is free in the bloodstream, however, it is found associated with HDL to maintain it inactive. This is because the triacylglycerides in HDL serve as a substrate, but the lipoprotein contains proteins around the triacylglycerides that can prevent the triacylglycerides from being broken down by HL.
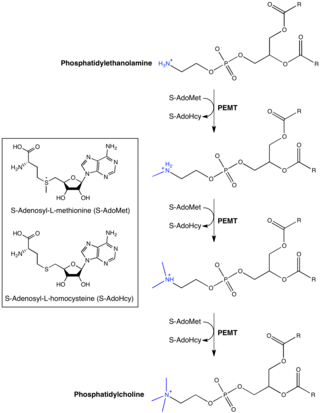
Phosphatidylethanolamine N-methyltransferase is a transferase enzyme which converts phosphatidylethanolamine (PE) to phosphatidylcholine (PC) in the liver. In humans it is encoded by the PEMT gene within the Smith–Magenis syndrome region on chromosome 17.
Apolipoprotein A-IV is plasma protein that is the product of the human gene APOA4.
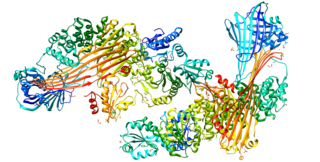
Microsomal triglyceride transfer protein large subunit is a protein that in humans is encoded by the MTTP, also known as MTP, gene.
ATP-binding cassette sub-family G member 8 is a protein that in humans is encoded by the ABCG8 gene.

Apolipoprotein M is an apolipoprotein and member of the lipocalin protein family that in humans is encoded by the APOM gene. It is found associated with high density lipoproteins and to a lesser extent with low density lipoproteins and triglyceride-rich lipoproteins. The encoded protein is secreted through the plasma membrane but remains membrane-bound, where it is involved in lipid transport. Two transcript variants encoding two different isoforms have been found for this gene, but only one of them has been fully characterized. It lacks an external amphipathic motif and is uniquely secreted to plasma without cleavage of its terminal signal peptide. The average molecular weight is 21253 Da, and the monoisotopic molecular weight is 21239 Da.
Apolipoprotein L1 is a protein that in humans is encoded by the APOL1 gene. Two transcript variants encoding two different isoforms have been found for this gene.

Familial hypertriglyceridemia is a genetic disorder characterized by the liver overproducing very-low-density lipoproteins (VLDL). As a result, an affected individual will have an excessive number of VLDL and triglycerides on a lipid profile. This genetic disorder usually follows an autosomal dominant inheritance pattern. The disorder presents clinically in patients with mild to moderate elevations in triglyceride levels. Familial hypertriglyceridemia is typically associated with other co-morbid conditions such as hypertension, obesity, and hyperglycemia. Individuals with the disorder are mostly heterozygous in an inactivating mutation of the gene encoding for lipoprotein lipase (LPL). This sole mutation can markedly elevate serum triglyceride levels. However, when combined with other medications or pathologies it can further elevate serum triglyceride levels to pathologic levels. Substantial increases in serum triglyceride levels can lead to certain clinical signs and the development of acute pancreatitis.





