Кора головного мозку
Кора́ головно́го мо́зку — зовнішній шар нервової тканини головного мозку людини й інших видів ссавців. Кора головного мозку повздовжною щілиною (лат. fissura longitudinalis) поділена на дві великих частини, які називаються півкулями мозку або гемісферами — правою й лівою. Обидві гемісфери з'єднані знизу мозолистим тілом (лат. corpus callosum). Кора головного мозку відіграє ключову роль у виконанні мозком таких функцій як пам'ять, увага, сприйняття, мислення, мова, свідомість.
| Кора головного мозку | |
|---|---|
 Зовнішній шар кори виділений фіолетовим. | |
 Нейрони кори головного мозку, зафарбовані методом Гольджі | |
| Деталі | |
| Частина від | Головний мозок людини |
| Ідентифікатори | |
| Латина | Cortex cerebri |
| MeSH | D002540 |
| NeuroNames | 39 |
| NeuroLex ID | birnlex_1494 |
| TA98 | A14.1.09.003 і A14.1.09.301 |
| TA2 | 5527 і 5528 |
| FMA | 61830 |
| Анатомічна термінологія | |
У великих ссавців кора головного мозку збирається в брижі, що дає більшу площу її поверхні в тому самому об'ємі черепа. Брижі називаються звивинами, а між ними пролягають борозни, й глибші — щілини. Головний мозок людини на дві третини ховається у борознах і щілинах. Кора головного мозку має товщину від 2 до 4 мм.[1]
Кора утворена сірою речовиною, яка складається переважно з тіл клітин, переважно, астроцитів, та капілярів. Тому навіть візуально тканина кори відрізняється від білої речовини, яка залягає глибше й складається переважно з білих мієлінових волокон — аксонів нейронів.
Зовнішня частина кори, так званий неокортекс (лат. neocortex), найбільш еволюційно молода частина кори у ссавців, має до шести клітинних шарів. Нейрони різних шарів поєднані між собою в кортикальні мініколонки. Різні ділянки кори, знані як поля Бродмана, різняться між собою за цитоархітектонікою (гістологічною структурою) й функціональною роллю в чутливості, мисленні, свідомості й пізнанні.[2][3]
Розвиток
ред.
Кора головного мозку розвивається з ембріональної ектодерми, а саме, з передньої частини нервової пластинки[4][5]. [31] Нервова пластинка згортається і формує нервову трубку. З порожнини всередині нервової трубки виникає система шлуночків, а з епітеліальних клітин її стінок — нейрони й глія. З фронтальної частини нервової пластинки формується передній мозок, великі півкулі головного мозку і потім — кора [5][6]
Зона росту кортикальних нейронів, так звана зона «S» знаходиться поруч із системою шлуночків головного мозку. Ця зона містить клітини-попередники, які пізніше в процесі діференціації стають гліальними клітинами і нейронами.[7] Гліальні волокна, утворені в перших діленнях клітин-попередників, радіально орієнтовані, охоплюють товщину кори з шлуночкової зони до м'якої мозкової оболони (лат. Pia mater) й утворюють «рейки» для міграції нейронів назовні від шлуночкової зони.[8][9][10] Ці дочірні нервові клітини стають пірамідними клітинами кори.[11] Процес розвитку чітко регламентований в часі й керується сотнями генів і механізмами енергорегуляції.[12] В процесі розвитку формується й пошарова структура кори.[13][14][15]
Клітинні шари
ред.


Кожен з клітинних шарів має характерну щільність нервових клітин і зв'язків з іншими ділянками. Існують прямі зв'язки між різними ділянками кори й непрямі зв'язки, наприклад, через таламус. Один з типових зразків кортикального розшарування — смужка Дженнарі в первинній зоровій корі. Це тяж візуально білішої тканини, помітний неозброєним оком в основі шпорної борозни (лат. sulcus calcarinus) в потиличній долі (лат. lobus occipitalis). Смужка Дженнарі складається з аксонів, які несуть візуальну інформацію з таламуса в четвертий шар зорової кори.
Зафарбування колонок клітин та їхніх аксонів дозволило нейроанатомам початку ХХ ст. зробити детальний опис пошарової структури кори у різних видів. Після робіт Корбініана Бродмана (1909) нейрони в корі були згруповані в шість основних шарів — від зовнішніх, які прилягають до м'якої мозкової оболони; до внутрішніх, що межують з білою речовиною:
- Шар I, молекулярний шар, містить кілька розрізнених нейронів і складається переважно з вертикально (апікально) орієнтованих дендритів пірамідних нейронів і горизонтально орієнтованих аксонів, та гліальних клітин.[16] Протягом розвитку в цьому шарі присутні клітини Кахаля-Ретціуса[17] та субпіальні клітини (клітини, що знаходяться одразу під (м'якою мозковою оболоною — лат. pia mater) зернистого шару[18]. Також тут іноді зустрічаються шипуваті астроцити. Апікальні пучки дендритів, як вважається, мають велике значення для реципрокних з'єднань («зворотного зв'язку») в корі головного мозку, й беруть участь у виконанні функцій асоціативного навчання й уваги.[19][20][21][22]
- Шар II, зовнішній гранулярний шар, містить малі пірамідні нейрони й численні зірчасті нейрони (дендрити яких виходять з різних боків тіла клітини, утворюючи форму зірки).
- Шар III, зовнішній пірамідний шар, містить переважно малі й середні пірамідні й непірамідні нейрони з вертикально орієнтованими інтракортикальними (тими, які в межах кори). Клітинні шари з І по ІІІ — головні мішені внутрішньопівкульних аферентів, а ІІІ-й шар — головне джерело кортико-кортикальних еферентів.
- Шар IV, внутрішній гранулярний шар, містить різні типи пірамідних і зірчастих нейронів і слугує головною мішенню таламокортикальних (від таламуса до кори) аферентних волокон.
- Шар V, внутрішній пірамідний шар, містить великі пірамідні нейрони, аксони яких залишають кору й прямують до підкіркових структур (таких як базальні ганглії). У первинній моторній корі цей шар містить клітини Беца, аксони яких йдуть через внутрішню капсулу, стовбур мозку та спинний мозок і формують кортикоспінальний шлях, який здійснює контроль довільних рухів.
- Шар VI, поліморфний або мультиформний шар, містить трохи пірамідних нейронів і багато поліморфних нейронів ; еферентні волокна з цього шару йдуть до таламуса, встановлюючи зворотний (реципрокний) зв'язок між таламусом і корою.[23][24][24].
Кортикальні шари не просто складовані один на один. Існують характерні зв'язки між різними шарами й типами клітин у них, які пронизують усю товщу кори. Базовою функціональною одиницею кори вважається кортикальна мініколонка (вертикальна колонка нейронів в корі головного мозку, яка проходить через її шари. Мініколонка включає від 80 до 120 нейронів в усіх зонах мозку, окрім первинної зорової кори приматів)[25][26].
Ділянки кори без четвертого (внутрішнього гранулярного) шару називаються агранулярними, з рудиментарним гранулярним шаром — дизгранулярними.[27] Швидкість обробки інформації в межах кожного шару різна. Так у ІІ і ІІІ — повільна, з частотою (2 Hz) у той час коли в частота осціляції в шарі V набагато швидше — 10–15 Hz.[28]
Зони кори
ред.Анатомічно кора може бути поділена на чотири частки, які мають назви відповідні до назв кісток черепа, що їх прикривають:
- Лобова частка (мозок), (лат. lobus frontalis)
- Скронева частка, (лат. lobus temporalis)
- Тім'яна частка, (лат. lobus parietalis)
- Потилична частка, (лат. lobus occipitalis)

-
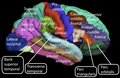 Латеральна (зовнішньо-бічна) поверхня кори головного мозку
Латеральна (зовнішньо-бічна) поверхня кори головного мозку -
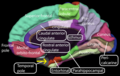 Медіальна (внутрішньо-бічна) поверхня кори головного мозку
Медіальна (внутрішньо-бічна) поверхня кори головного мозку


З огляду на особливості ламінарної (пошарової) структури, кора поділяється на неокортекс, і алокортекс:
- Неокортекс (лат. neocortex; інші назви — ізокортекс, лат. isocortex та неопалліум, лат. neopallium) — частина зрілої кори головного мозку з шістьма клітинними шарами. Зразками неокортикальних ділянок — це Поле Бродмана 4, також знане як первинна моторна кора, первинна зорова кора, або поле Бродмана 17. Неокортекс поділяється на два типи: ізокортекс (справжній неокортекс, зразки котрого, поля Бродмана 24,25 і 32 щойно розглянуті) і проізокортекс, який представляють, зокрема, поле Бродмана 24, поле Бродмана 25 і поле Бродмана 32
- Алокортекс (лат. allocortex) — частина кори з кількістю клітинних шарів менше шести, теж поділяється на частини: палеокортекс (лат. paleocortex) з тришаровою, архікортекс (лат. archicortex) з чотирьох-п'ятьох, та прилеглий до них періалокортекс (лат. periallocortex). Прикладами ділянок з такою пошаровою структурою є нюхова кора: склепінчаста звивина (лат. gyrus fornicatus) з крючком (лат. uncus), гіпокамп (лат. hippocampus) й ближні до нього структури.
Існує й «перехідна» (між алокортексом і неокортексом) кора, яка має назву паралімбічної, де клітинні шари 2,3 та 4 зливаються. Ця зона містить проізокортекс (з неокортексу) і періалокортекс (з алокортексу).
Поля Бродмана
ред.
Різні ділянки кори залучені до виконання різних функцій. Побачити й зафіксувати цю різницю можна в різний спосіб — досліджуючи ураження певних ділянок, порівнюючи патерни електричної активності, використовуючи методики нейровізуалізації, вивчаючи клітинну структуру. На основі таких відмінностей дослідники класифікують ділянки кори.
Найбільш відомою й цітованою вже протягом століття є класифікація, яку створив у 1905—1909 рр німецький дослідник Корбініан Бродман. Він поділив кору головного мозку на 51 ділянку на основі цитоархітектоніки нейронів, який він вивчав у корі головного мозку за допомогою фарбування клітин за Ніслем. Бродман опублікував свої карти областей кори головного мозку у людини, мавп та інших видів у 1909 році[29].
Поля Бродмана активно й докладно обговорюються, дискутуються, уточнюються, і перейменуються протягом майже століття й залишаються найбільш широко відомими і часто цитованими структурами цитоархітектонічної організації кори головного мозку людини.
Багато з полів Бродмана, початково визначені виключно за їхньою нейрональною організацією, пізніше були асоційовані відповідно до кореляції з різними корковими функціями. Наприклад, Поля 3, 1 & 2 — первинна соматосенсорна кора; поле 4 є первинною моторною корою; поле 17 є первинною зоровою корою, а поля поля 41 і 42 найбільше корелюють із первинною слуховою корою. Визначення відповідності процесів Вищої нервової діяльності до ділянок кори головного мозку й прив'язка до конкретних полів Бродмана здійснюється за допомогою нейрофізіологічних досліджень, функціональної магнітнорезонансної томографії та інших методик (так як це було, наприклад, зроблено з прив'язкою зон Брока мовлення й мови до полів Бродмана 44 і 45). Однак, за допомогою функціональної візуалізації можна тільки приблизно визначити локалізацію активації мозкових процесів у полях Бродмана. А для точного визначення їхніх меж в кожному окремому мозку потрібно гістологічне дослідження.[30]
Товщина кори
ред.У ссавців, видів з більшими розмірами мозку (в абсолютному вираженні, а не тільки по відношенню до розміру тіла), кора, як правило, має більшу товщину кору[31] . Діапазон, однак, не дуже великий. Найменші ссавці, такі як землерийки, мають товщину неокортексу приблизно 0,5 мм; а види з найбільшим мозком, такі як люди і китоподібні, мають товщину 2,3-2,8 мм. Існує приблизно логарифмічна залежність між вагою мозку і товщиною кори.[31]
Магніторезонансна томографія (МРТ) мозку робить можливими прижиттєві заміри товщини кори й порівнняння відносно до розмірів тіла. Товщина різних ділянок варіативна, але в цілому, сенсорні (чутливі) ділянки кори тонші за моторні (рухові).[32] В одному з досліджень показано залежність товщини кори від рівня інтелекту.[33] Інше дослідження показало більшу товщину кори осіб, що страждають на мігрень.[34][35] Щоправда, інші дослідження показують відсутність такого зв'язку.[36][37]
Звивини, борозни й щілини
ред.Разом ці три елементи — звивини, борозни й щілини, створюють велику площу поверхні мозку людини та інших ссавців. При погляді на людський мозок, помітно, що дві третини поверхні приховані в пазах. Як борозни, так і щілини являють собою заглиблення в корі, але вони розрізняються за розміром. Борозна-це неглибокий паз, який оточує звивини. Щілина — це велика борозна, яка ділить мозок на частки, а також на дві півкулі як, наприклад медіальна поздовжня щілина[38]. Однак ця відмінність не завжди чітка. Наприклад, латеральна борозна також знана як бічна щілина і як «Сільвієва борозна» і «центральна борозна», також знана як Центральна щілина і як «Роландова борозна».
Це дуже важливо в умовах, коли розмір мозку обмежується внутрішнім розміром черепа. Збільшення поверхні кори головного мозку за допомогою системи звивин і борозен збільшує кількість клітин, які беруть участь у виконанні мозком таких функцій як пам'ять,увага, сприйняття, мислення, мова, свідомість.[39][40] [41]
Кровопостачання
ред.Постачання артеріальної крові до головного мозку й кори, зокрема, відбувається з двох артеріальних басейнів — внутрішньої сонної і хребетної артерії. Кінцевий відділ внутрішньої сонної артерії розгалужується на гілки — передню мозкову й середню мозкову артерію. У нижніх (базальних) відділах мозку артерії утворюють Вілізієве коло, завдяки котрому відбувається перерозподіл артеріальної крові між артеріальними басейнами.
-
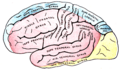 Зовнішня поверхня головного мозку на якій позначені ділянки, що кровопостачаються мозковими артеріями. Ділянка позначена синім кольором відповідає передній мозковій артерії. Ділянка задньої мозкової артерії позначена жовтим
Зовнішня поверхня головного мозку на якій позначені ділянки, що кровопостачаються мозковими артеріями. Ділянка позначена синім кольором відповідає передній мозковій артерії. Ділянка задньої мозкової артерії позначена жовтим -
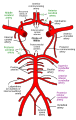 Схема Вілізієвого кола та артерій головного мозку: Передня мозкова артерія - англ. Anterior cerebral artery; Середня мозкова артерія - англ. Middle cerebral artery; Задня мозкова артерія - англ. Posterior cerebral artery; Хребетна артерія - англ. Vertebral artery
Схема Вілізієвого кола та артерій головного мозку: Передня мозкова артерія - англ. Anterior cerebral artery; Середня мозкова артерія - англ. Middle cerebral artery; Задня мозкова артерія - англ. Posterior cerebral artery; Хребетна артерія - англ. Vertebral artery


Середня мозкова артерія
ред.Середня мозкова артерія (лат. a. cerebri media) є найбільшою гілкою внутрішньої сонної артерії.[42][3] Порушення кровообігу в ній може призводити до розвитку ішемічного інсульту та синдрому середньої мозкової артерії з наступними симптомами:
- Параліч (плегія) або парез протилежних до ураження м'язів обличчя та руки
- Втрата сенсорної чутливості протилежних до ураження м'язів обличчя та руки
- Ураження домінантної півкулі (часто лівої) головного мозку та розвитку афазії Брока або афазії Верніке
- Ураження недомінантної півкулі (часто правої) головного мозку призводить до односторонньої просторової агнозії з протилежного до ураження боку
- Інфаркти в зоні середньої мозкової артерії призводять до déviation conjuguée, коли зіниці очей рухаються в бік сторони ураження головного мозку.
Передня мозкова артерія
ред.Передня мозкова артерія — менша гілка внутрішньої сонної артерії. Досягнувши медіальної поверхні півкуль головного мозку, передня мозкова артерія йде до потиличної частки. Вона кровопостачає медіальні ділянки півкуль до рівня тім'яно-потиличної борозни, ділянку верхньої лобної звивини, ділянку тім'яної частки, а також ділянки нижніх медіальних відділів очноямкових звивин. Симптоми її ураження:
- Парез ноги або геміпарез з переважним ураженням ноги на протилежному боці.
- Закупорка парацентральної гілки зумовлює монопарез стопи, який нагадує периферичний парез. Можуть спостерігатися затримка або нетримання сечі. З'являються рефлекси орального автоматизму та хватальні феномени, патологічні стопні згинальні рефлекси: Россолімо, Бехтерева, Жуковського. Виникають зміни психічного стану, зумовлені ураженням лобової частки: зниження критики, пам'яті, немотивована поведінка.
Задня мозкова артерія
ред.Задня мозкова артерія — парна судина, яка кровопостачає задні відділи мозку (потиличну частку). Має анастомоз із середньою мозковою артерією Її ураження призводять до:
- гомонімна (чи верхньоквадрантна) геміанопсія (випадіння частки поля зору)
- метаморфопсія (порушення зорового сприйняття величини або форми предметів і простору) і зорова агнозія,
- Алексія,
- Сенсорна афазія,
- Транзиторні (минучі) амнезії;
- Трубчастий зір,
- Кіркова сліпота (при збереженні реакції на світло),
- Прозопагнозія,
- Порушення орієнтації в просторі
- Втрата топографічної пам'яті
- Набута ахроматопсія — недостатність колірного зору
- Синдром Корсакова (порушення оперативної пам'яті)
- емоційно-афективні порушення
Див. також
ред.Примітки
ред.- ↑ Kandel, Eric R.; Schwartz, James H.; Jessell, Thomas M. (2000). Principles of Neural Science (вид. Fourth). United State of America: McGraw-Hill. с. 324. ISBN 0-8385-7701-6.
- ↑ Неттер Ф. (2004). Атлас анатомії людини (переклад Цегельського А.А.). Львів: Наутілус. с. 592. ISBN 966-95745-8-7.
- ↑ а б Human Anatomy & Physiology (вид. 7th). Benjamin Cummings. 2006. ISBN 978-0805359091.
- ↑ Pletikos, Mihovil; Sousa, Andre MM (22 січня 2014). Temporal Specification and Bilaterality of Human Neocortical Topographic Gene Expression. Neuron. 81 (2): 321—332. doi:10.1016/j.neuron.2013.11.018. PMC 3931000. PMID 24373884. Архів оригіналу за 24 вересня 2015. Процитовано 6 квітня 2017.
- ↑ а б Natasha Warren; Damira Caric; Thomas Pratt; Julia A. Clausen; Pundit Asavaritikrai; John O. Mason; Robert E. Hill; David J. Price; Oxford Journals (1999). The transcription factor, Pax6, is required for cell proliferation and differentiation in the developing cerebral cortex. National Institutes of Health. с. 627—635. PMID 10498281.
{{cite web}}: Пропущений або порожній|url=(довідка) - ↑ Larsen, W J. Human Embryology 3rd edition 2001. pp 421—422 ISBN 0-443-06583-7
- ↑ Stephen C. Noctor; Alexander C. Flint; Tamily A. Weissman; Ryan S. Dammerman; Arnold R. Kriegstein (2001). Neurons derived from radial glial cells establish radial units in neocortex. Nature (journal). 409 (6821): 714—720. doi:10.1038/35055553. PMID 11217860. Архів оригіналу за 29 вересня 2007. Процитовано 6 квітня 2017.
- ↑ Rakic, P (October 2009). Evolution of the neocortex: a perspective from developmental biology. Nature reviews. Neuroscience. 10 (10): 724—35. doi:10.1038/nrn2719. PMC 2913577. PMID 19763105.
- ↑ Rakic, P (November 1972). Extrinsic cytological determinants of basket and stellate cell dendritic pattern in the cerebellar molecular layer. The Journal of Comparative Neurology. 146 (3): 335—54. doi:10.1002/cne.901460304. PMID 4628749.
- ↑ Calegari, F; Haubensack W; Haffner C; Huttner WB (2005). Selective lengthening of the cell cycle in the neurogenic subpopulation of neural progenitor cells during mouse brain development. J Neurosci. 25 (28): 6533—8. doi:10.1523/jneurosci.0778-05.2005. PMID 16014714.
- ↑ P. Rakic (1988). Specification of cerebral cortical areas. Science (journal). 241 (4862): 170—176. doi:10.1126/science.3291116. PMID 3291116. Архів оригіналу за 30 вересня 2007. Процитовано 6 квітня 2017.
{{cite journal}}: Текст «Science» проігноровано (довідка) - ↑ Hu, X.L.; Wang,Y.; Shen, Q. (2012). Epigenetic control on cell fate choice in neural stem cells. Protein & Cell. 3 (4): 278—290. doi:10.1007/s13238-012-2916-6. PMC 4729703. PMID 22549586.
- ↑ Kostović, Ivica. Developmental history of the transient subplate zone in the visual and somatosensory cortex of the macaque monkey and human brain. Journal of Comparative Neurology. 297 (3): 441—470. doi:10.1002/cne.902970309. Архів оригіналу за 24 вересня 2015. Процитовано 6 квітня 2017.
- ↑ Fukuchi-Shimogori, T; Grove, EA (2 листопада 2001). Neocortex patterning by the secreted signaling molecule FGF8. Science. 294 (5544): 1071—4. doi:10.1126/science.1064252. PMID 11567107.
- ↑ Garel, S; Huffman, KJ; Rubenstein, JL (May 2003). Molecular regionalization of the neocortex is disrupted in Fgf8 hypomorphic mutants. Development (Cambridge, England). 130 (9): 1903—14. doi:10.1242/dev.00416. PMID 12642494.
- ↑ Shipp, Stewart (17 червня 2007). Structure and function of the cerebral cortex. Current Biology. 17 (12): R443—9. doi:10.1016/j.cub.2007.03.044. PMID 17580069. Архів оригіналу за 2 жовтня 2010. Процитовано 17 лютого 2009.
- ↑ Meyer, Gundela; Goffinet, André M.; Fairén, Alfonso (1999). Feature Article: What is a Cajal–Retzius cell? A Reassessment of a Classical Cell Type Based on Recent Observations in the Developing Neocortex. Cereb. Cortex. 9 (8): 765—775. doi:10.1093/cercor/9.8.765. PMID 10600995. Архів оригіналу за 21 лютого 2015. Процитовано 4 квітня 2017.
- ↑ Judaš, Miloš; Pletikos, Mihovil (2010). The discovery of the subpial granular layer in the human cerebral cortex. Translational Neuroscience. 1 (3): 255—260. doi:10.2478/v10134-010-0037-4. Архів оригіналу за 12 березня 2016. Процитовано 4 квітня 2017.
- ↑ Gilbert CD, Sigman M (2007). Brain states: top-down influences in sensory processing. Neuron. 54 (5): 677—96. doi:10.1016/j.neuron.2007.05.019. PMID 17553419.
- ↑ Cauller L (1995). Layer I of primary sensory neocortex: where top-down converges upon bottom-up. Behav Brain Res. 71 (1–2): 163—70. doi:10.1016/0166-4328(95)00032-1. PMID 8747184.
- ↑ Rubio-Garrido P, Pérez-de-Manzo F, Porrero C, Galazo MJ, Clascá F (2009). Thalamic input to distal apical dendrites in neocortical layer 1 is massive and highly convergent. Cereb Cortex. 19 (10): 2380—95. doi:10.1093/cercor/bhn259. PMID 19188274.
- ↑ Jones EG (1998). Viewpoint: the core and matrix of thalamic organization. Neuroscience. 85 (2): 331—45. doi:10.1016/S0306-4522(97)00581-2. PMID 9622234.
- ↑ Creutzfeldt, O. 1995. Cortex Cerebri. Springer-Verlag.
- ↑ а б Lam YW, Sherman SM (2010). Functional Organization of the Somatosensory Cortical Layer 6 Feedback to the Thalamus. Cereb Cortex. 20 (1): 13—24. doi:10.1093/cercor/bhp077. PMC 2792186. PMID 19447861.
- ↑ Mountcastle V (1997). The columnar organization of the neocortex. Brain. 120 (4): 701—722. doi:10.1093/brain/120.4.701. PMID 9153131.
- ↑ HUBEL DH, WIESEL TN (October 1959). Receptive fields of single neurones in the cat's striate cortex. J. Physiol. (Lond.). 148 (3): 574—91. doi:10.1113/jphysiol.1959.sp006308. PMC 1363130. PMID 14403679.
- ↑ S.M. Dombrowski, C.C. Hilgetag, and H. Barbas. Quantitative Architecture Distinguishes Prefrontal Cortical Systems in the Rhesus Monkey [Архівовано 29 серпня 2008 у Wayback Machine.].Cereb. Cortex 11: 975—988. «…they either lack (agranular) or have only a rudimentary granular layer IV (dysgranular).»
- ↑ Sun W, Dan Y (2009). Layer-specific network oscillation and spatiotemporal receptive field in the visual cortex. Proc Natl Acad Sci U S A. 106 (42): 17986—17991. doi:10.1073/pnas.0903962106. PMC 2764922. PMID 19805197.
- ↑ Brodmann K (1909). Vergleichende Lokalisationslehre der Grosshirnrinde (нім.). Leipzig: Johann Ambrosius Barth. Архів оригіналу за 29 листопада 2014. Процитовано 4 квітня 2017.
- ↑ Principles of Anatomy and Physiology 12th Edition — Tortora, Page 519-fig. (14.15)
- ↑ а б Nieuwenhuys R, Donkelaar HJ, Nicholson C (1998). The central nervous system of vertebrates, Volume 1. Springer. с. 2011—2012. ISBN 978-3-540-56013-5.
- ↑ Frithjof Kruggel; Martina K. Brückner; Thomas Arendt; Christopher J. Wiggins; D. Yves von Cramon (2003). Analyzing the neocortical fine-structure. Medical Image Analysis. 7 (3): 251—264. doi:10.1016/S1361-8415(03)00006-9.
- ↑ Katherine L. Narr; Roger P. Woods; Paul M. Thompson; Philip Szeszko; Delbert Robinson; Teodora Dimtcheva; Mala Gurbani; Arthur W. Toga; Robert M. Bilder (2007). Relationships between IQ and Regional Cortical Grey Matter Thickness in Healthy Adults. Cerebral Cortex. 17 (9): 2163—2171. doi:10.1093/cercor/bhl125. PMID 17118969.
- ↑ Alexandre F.M. DaSilva; Cristina Granziera; Josh Snyder; Nouchine Hadjikhani (2007). Thickening in the somatosensory cortex of patients with migraine. Neurology. 69 (21): 1990—1995. doi:10.1212/01.wnl.0000291618.32247.2d. PMID 18025393.
- ↑ Catharine Paddock (20 листопада 2007). Migraine Sufferers Have Thicker Brain Cortex. Medical News Today. Архів оригіналу за 11 травня 2008. Процитовано 5 квітня 2017.
- ↑ Datte R, Detre JA (Oct 2011). Absence of changes in cortical thickness in patients with migraine. Cephalagia. 31 (14): 1452—8. doi:10.1177/0333102411421025. PMC 3512201. PMID 21911412.
- ↑ Habib M (2000). The neurological basis of developmental dyslexia: an overview and working hypothesis. Brain. 123 (12): 2373—99. doi:10.1093/brain/123.12.2373. PMID 11099442.
- ↑ Carlson, N. R. (2013). Physiology of Behavior. Upper Saddle River, NJ: Pearson Education Inc.
- ↑ Cusack, R. (2005). The intraparietal sulcus and perceptual organization. Journal of Cognitive Neuroscience, 17(4), 641—651. doi: 10.1162/0898929053467541
- ↑ Ono, Kubick, Abernathey, Atlas of the Cerebral Sulci, Thieme Medical Publishers, 1990. ISBN 0-86577-362-9. ISBN 3-13-732101-8.
- ↑ Toro, Roberto; Perron, Michel; Pike, Bruce; Richer, Louis; Veillette, Suzanne; Pausova, Zdenka; Paus, Tomáš (1 жовтня 2008). Brain Size and Folding of the Human Cerebral Cortex. Cerebral Cortex (англ.). 18 (10): 2352—2357. doi:10.1093/cercor/bhm261. ISSN 1047-3211. PMID 18267953.
- ↑ Неттер Ф. (2004). Атлас анатомії людини (переклад Цегельского А.А.). Львів: Наутілус. с. 592. ISBN 966-95745-8-7.
Посилання
ред.- Більш детально про порушення кровообігу в різних басейнах [Архівовано 6 квітня 2017 у Wayback Machine.]
- Більш детально про кровопостачання головного й спинного мозку [Архівовано 6 квітня 2017 у Wayback Machine.]