

Abstract
Skin morphogenesis occurs in successive stages. First, the skin forms distinct regions (macropatterning). Then skin appendages with particular shapes and sizes form within each region (micropatterning). Ectopic DKK expression inhibited dermis formation in feather tracts and individual buds, implying the importance of Wnts, and prompted the assessment of individual Wnt functions at different morphogenetic levels using the feather model. Wnt 1, 3a, 5a and 11 initially were expressed moderately throughout the feather tract then were up-regulated in restricted regions following two modes: Wnt 1 and 3a became restricted to the placodal epithelium, then to the elongated distal bud epidermis; Wnt 5a and 11 intensified in the inter-tract region and interprimordia epidermis or dermis, respectively, then appeared in the elongated distal bud dermis. Their role in feather tract formation was determined using RCAS mediated misexpression in ovo at E2/E3. Their function in periodic feather patterning was examined by misexpression in vitro using reconstituted E7 skin explant cultures. Wnt 1 reduced spinal tract size, but enhanced feather primordia size. Wnt 3a increased dermal thickness, expanded the spinal tract size, reduced interbud domain spacing, and produced non-tapering ‘giant buds’. Wnt 11 and dominant negative Wnt 1 enhanced interbud spacing, and generated thinner buds. In cultured dermal fibroblasts, Wnt 1 and 3a stimulated cell proliferation and activated the canonical beta-catenin pathway. Wnt 11 inhibited proliferation but stimulated migration. Wnt 5a and 11 triggered the JNK pathway. Thus distinctive Wnts have positive and negative roles in forming the dermis, tracts, interbud spacing and the growth and shaping of individual buds.
Keywords: Beta-catenin, Pattern formation, Feather morphogenesis, Organogenesis, Hair
1. Introduction
Skin appendage formation results from a series of interactions between the mesenchyme (dermis) and epidermis. As tissue morphogenesis and differentiation progress, the nature of these signals and their interpretation by the receiving tissue are altered through hierarchical morphogenesis. The dermal precursor cells initially migrate from the dermomyotome to the subectoderm region. The presumptive dermis then forms distinct skin regions. Skin appendage primordia are induced within the skin regions as epidermal placodes and dermal condensations. Finally, each skin appendage grows with a characteristic size and shape. These processes all take place in the chicken integument, which is easily accessible to experimentation. Therefore, the chicken provides an excellent research model to study hierarchical morphogenesis during skin formation from the dermatome to individual skin appendages (Chuong, 1993; Chuong et al.,2000; Chen and Chuong, 1999; Prum and Dyck, 2003). In this paper, we primarily focus on feathers that form within the chicken dorsal tract.
Many signaling pathways, such as Wnt, FGF, BMP, SHH, etc. are used repetitively to regulate tissue interactions in different cellular contexts and at different developmental stages. Here we focus on the Wnt signaling cascade. Wnt molecules are potent developmental morphogens that specify different cell fates, provide positional information, organize pattern formation, and regulate cell migration, proliferation and apoptosis throughout embryogenesis (Wodarz and Nusse, 1998; Dierick and Bejsovec, 1999). Humans express 19 known Wnts and about 10 frizzled (Fz) receptors (Malbon et al., 2001). Wnts can trigger at least three downstream pathways: in the canonical Wnt 1/wingless pathway, cytoplasmic beta-catenin accumulates, translocates to the nucleus and activates the transcription of T cell factor (TCF) dependent target genes (Lustig and Behrens, 2003); the Wnt/Ca+2 pathway acts by stimulating G protein dependent intracellular calcium release and activating protein kinase C (PKC) and calmodulin-dependent protein kinase II (Kühl et al., 2000); and the planar cell polarity pathway activates c-Jun N-terminal kinase (JNK) (Weston and Davis, 2002).
What is known about the role of Wnts in chicken skin morphogenesis? At the onset of chicken skin formation, dermal precursors migrate to the dorsal trunk from the medial dermomyotome of the somites at around E3 (Mauger, 1972; Olivera-Martinez et al., 2002). These cells express Wnt 11 (Tanda et al., 1995). The survival and migration of these dermal precursors require signals derived from the dorsal neural tube and Wnt 1 could substitute for the dorsal neural tube signals and induce Wnt 11 expression in the migrating cells. (Olivera-Martinez et al., 2001).
In the subsequent macropatterning process, the developing skin establishes several skin regions including the spinal, scapular, femoral, caudal tracts, etc. where the epithelium and mesenchyme become competent to execute micropatterning, the induction of individual feather primordia (Sengel, 1976). The regions between these tracts, called apteria, form few feathers. The origin of the dermis influences regional specificity (Dhouailly, 1978). The dorsal spinal tract begins from the midline region above the spinal cord around stage 26-27 (E5) and then expands bilaterally, characterized by the formation of a dense dermis. Both cell migration and proliferation contribute to increase the cell density in the tract region (Chuong and Widelitz, 1998). Wnt pathway modulation has profound effects on skin patterning. The loss of mammary glands, teeth and skin appendages in transgenic mice over-expressing Dickkopf 1 (DKK1), a potent diffusible Wnt inhibitor (Andl et al., 2002), suggests that Wnts are involved in regulating the response of skin regions.
In each tract, the homogeneous dense dermis and the overlying epidermis are transformed into periodically arranged feather primordia (Chuong and Edelman, 1985), which appear in a temporal and spatial sequence (Mayerson and Fallon, 1984). They first propagate linearly to establish the primary row and then bilaterally (in the spinal tract) or unilaterally (in the scapular or femoral tract) to the tract margin to set up the micropatterns. The dermis, rather than the epidermis, is the main factor determining skin appendage size and spacing (Jiang et al., 1999). Therefore, whether a region of epidermis becomes part of the tract or apteria results from differential dermis development. The forced expression of exogenous beta-catenin in chick embryos induced ectopic feathers in scale forming and apteric regions (Noramly et al., 1999; Widelitz et al., 2000). Lef1 is expressed in the dermis of mouse vibrissae and is required for initial vibrissae formation (Kratochwil et al., 1996). Wnt 10a and 10b are up-regulated and Wnt 5a is expressed in developing hair dermal condensations. These studies suggest that the Wnt pathway is involved in initial skin appendage induction.
Following induction, feather buds establish anterior– posterior asymmetry, then proximal–distal asymmetry. They then invaginate to form feather follicles. Wnt signaling at each step influences feather morphogenesis. Within individual skin appendages, Wnts aid in forming localized growth zones that shape the developing feathers. Wnt 6 over expression induced localized outgrowths from different feather regions (Chodankar et al., 2003). Wnt 7a localizes to the posterior feather epidermis (Chuong et al., 1996). Ectopic Wnt 7a expression produced ‘posteriorized’ feather buds with a flattened, plateau shape containing widely dispersed proliferating cells (Widelitz et al., 1999). Ectopic Wnt 3 expression in the hair follicle outer root sheath causes hair shaft fragility and dysregulation of hair intermediate filament formation (Millar et al., 1999). These studies suggest that the Wnts also regulate epithelial appendage morphogenesis. At each morphogenetic level, cell proliferation, adhesion, migration and differentiation are differentially regulated in space and time.
The current work systematically explores the roles of Wnt signaling during different levels of chicken skin formation. In this study, we found that Dkk inhibited dermis formation when applied early in skin development and individual feather bud formation at later times. Wnt 1 inhibited dermis formation when misexpressed at the midline initiation stage, but enhanced feather formation when expressed later in development. Wnt 3a expanded the feather tract size and enhanced individual feather formation. Wnt 11 had no observable effect on skin regionalization, but inhibited individual feather formation and expanded interbud spacing. The potential mechanisms underlying these effects were characterized.
2. Results
2.1. Expression patterns of Wnts in embryonic chicken skin
To analyze possible Wnt signaling involvement in tract and subsequent feather formation, we performed whole mount and section in situ hybridizations on chick embryos using digoxigenin-labeled antisense RNA probes.
Wnt 1 first was expressed homogenously in the epidermis and then became enhanced at stage 29 (E6) along the dorsal midline of the lumbar region, corresponding to the primary feather row at stage 33 (Fig. 1, E7.5). Subsequently, expression increased in the bud domain and decreased in the interbud domain as individual buds formed, so older buds were surrounded by a clear halo with a complete loss of Wnt 1 expression in the interbud area (Fig. 1). Just prior to the appearance of morphological anterior– posterior and proximal–distal asymmetry, expression moved to the distal feather bud epithelium at late stage 34 (Fig. 1, E8.5).
Fig. 1.

Expression of Wnt 1, 3a, 5a and 11 during skin development. Wnt expression during chicken embryo feather formation. All views are of the dorsal tract unless otherwise noted. Wnt 1 demonstrated a restrictive to bud expression mode. Wnt 1 expressed in the dorsal midline as a continuous stripe (arrow) with lower expression elsewhere at stage 29 (E6) was then restricted to the bud epithelium surrounded by a clear halo at stage 33 (E7.5), and moved to the distal short buds and long buds at late stage 34 (E8.5). Wnt 3a generally followed a similar expression pattern, as a continuous stripe in the stage 29 (E6) dorsal midline epithelium (arrow), then disappearing from the interbud area and only staining the bud domain at stage 31 (E7). Densely stained morphogenetic zones besides the primary row appeared before the newly emerging rows. Wnt 3a was localized to the distal short buds at stage 34 (E8). Insets show cross sections. Wnt 11 demonstrated a restrictive to interbud expression mode. Wnt 11 initiated as a continuous stripe in the primary row and then disappeared from the bud domain leaving clear circles as seen in the stage 31 (E7) cervical and femoral tracts. As the feather rows propagated laterally (arrow indicates direction), Wnt 11 strongly stained the interbud area of newly formed feather rows, faded from the interbud area of older rows, and finally appeared within the mesenchyme of short buds. Stage 35 (E9) embryo side view, Wnt 11 stains the apteric zones between the femoral/spinal (yellow asterisk) and humeral/spinal tracts (green asterisk). Wnt 11 appeared as a transient ring surrounding the feather bud base, then stained throughout the mesenchyme before disappearing under the early bud placodes (stage 31 sections). It then concentrated in the interbud mesenchyme, was lost from the bud areas (stage 34 sections), and gradually appeared in the distal and posterior short and long bud mesenchyme at stage 35—late 35 (E9–9.5). Wnt 5a was expressed all over the epithelium first at stage 30 (E6.5), then transiently disappeared in the bud epithelium at stage 30 (E6.5) and reappeared in the bud again at stage 31 (E7). In the short bud, Wnt 5a was expressed in the anterior bud epithelium and posterior bud mesenchyme, and retained the early stage interbud epithelial staining at stage 34 (E8). As the buds matured, the epithelial staining decreased while the mesenchymal staining increased, shifting the expression pattern to the posterior bud domain at stage 35 (E9). Wnt 5a was concentrated as a ring around the base of developing asymmetric feather buds, and stained the distal mesenchyme of long buds (E9, stage 35). Size bar = 500 μm, Size bar for insets = 100 μm. H and H developmental staging is indicated. Schematic drawing summarizing the expression patterns of Wnt 1, 3a, 5a and 11.
Wnt 3a also was expressed throughout the ectoderm and then appeared as a linear stripe during primary row initiation at stage 29 (Fig. 1, E6). Wnt 3a then became concentrated in the bud domain and absent in the interbud domain. In the distal dorsal trunk, Wnt 3a was expressed in the newly emerging feather bud rows at stage 31 (Fig. 1, E7) and then moved to the distal feather bud domain during anterior– posterior axis formation at stage 34 (Fig. 1, E8). Section in situ hybridization revealed that both Wnt 1 and Wnt 3a were initially expressed throughout the ectoderm and then became restricted to the bud ectoderm (Fig 1, see insets) in overlapping expression patterns.
Wnt 11 was always expressed in the mesenchyme. It initiated as a broad stripe that disappeared in the developing bud primordia (Fig. 1, stage 31, 34). It became intensely expressed in the interbud and apteric zones (Fig. 1, stage 35 wholemount, stage 34 skin, stage 31, 34 section). During short bud formation, Wnt 11 gradually appeared within the bud (Fig. 1, stage 34, 35), and concentrated in the posteriordistal bud mesenchyme during A–P axis formation (Fig. 1, late stage 35 skin, late stage 35 section). Wnt 11 expression also intensified in interbud mesenchyme in a pattern complementary to, rather than overlapping with the Wnt 1 and Wnt 3a expression pattern.
Wnt 5a was initially expressed throughout the epithelium and became restricted to the interbud area during placode formation (Fig. 1, stage 29, 30 section). Wnt 5a remained only transiently in the interbud zone compared with Wnt 11 (Fig. 1, stage 31 wholemount) and quickly appeared in the bud epithelium and mesenchyme. Wnt 5a was expressed in the anterior bud epithelium and in the posterior bud mesenchyme (Fig. 1, stage 31, 34). Mesenchymal staining was up-regulated and epithelial staining was down-regulated during A–P axis formation at stage 35 (Fig. 1, E9). In whole mounts, Wnt 5a staining appeared in the center of early short buds (Fig. 1, stage 31), and gradually shifted to the posterior bud (Fig. 1, stage 34). During bud elongation, Wnt 5a was expressed in the distal mesenchyme (Fig. 1, stage 35 skin). A ring surrounding the bud base was observed. The Wnt 5a expression pattern overlapped partially with Wnt 1/Wnt 3a and partially with Wnt 11.
The endogenous Wnt expression patterns are summarized in a schematic drawing (Fig. 1).
2.2. Retroviral mediated mis-expression of Wnt genes
To test Wnt function in tract and feather bud formation, exogenous Wnts were expressed from a replication competent avian sarcoma virus (RCAS). RCAS virus was applied using two experimental models. (1) Injection into E2 and E3 (stage 12–20) embryos, at early dermis development stages allows us to assay for factors that modulate the size of feather tracts. (2) Transduction of reconstituted explants from E7 (stage 31) embryos allows us to assay for factors that modulate the formation of individual feather buds. This enables us to analyze individual feather formation independent of tract formation.
DKK
We first used the Wnt antagonist DKK 1 to neutralize physiological Wnt activity. In in vivo assays, embryos transduced at stage 19-20 (E3) were harvested at stage 38 (E12). Anti-retroviral staining (p27) to detect virus showed the infection was widespread in both the epidermis and dermis (examples shown in Fig. 3). RCAS-DKK caused an expansion of the apteric region compared to controls. When feather buds did form, the interbud domains also expanded and feathers were thin (100%, n = 12; Fig. 2A). Longitudinal sections showed not only the absence of feather buds, but also a very thin dermis (Fig. 2B). The muscle layer also appeared to be less developed.
Fig. 3.
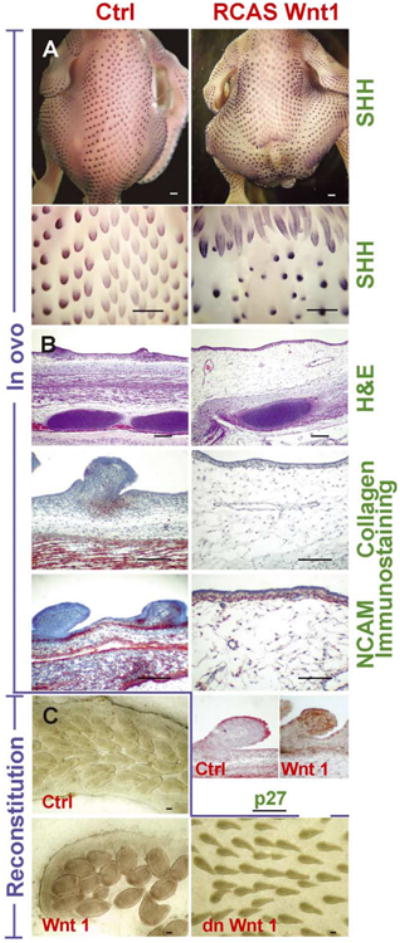
Dual functions of Wnt 1 in regulating the size of feather tracts and feather primordia. RCAS-Wnt 1 transduction in early embryos decreased both the tract and feather bud size. RCAS-Wnt 1 was injected to the body and amniotic cavity of stage 14 embryos. At stage 36 (E10), Shh staining revealed that the sacral and caudal parts of the spinal tract were blocked or prematurely truncated, allowing the femoral tracts on both sides to invade into this affected zone (A). The zone between the normal and affected spinal tract showed a depressed integument with inhibited feather development. A section through this region showed underdeveloped and uncondensed mesenchyme, loss of the deep dermis layer and thinning of the superficial dermis, compared with controls (B). Immunostaining showed loss of collagen II formation in the mesenchyme, compared with controls (B). Mesenchymal NCAM expression levels were decreased, compared with controls (B). The presence of retrovirus was confirmed in RCAS control and Wnt 1 transduced embryos by staining for p27 (B). In vitro reconstituted explants transduced with RCAS-Wnt 1 had increased bud size, compared to RCAS-LacZ infected controls (C), while incubation in conditioned medium from a dominant negative Wnt 1 secreting cell line produced thinner and sparser feather buds (C). Transduction was confirmed by staining for retroviral p27. Size bars; wholemount = 500 μm, sections and explants = 200 μm.
Fig. 2.
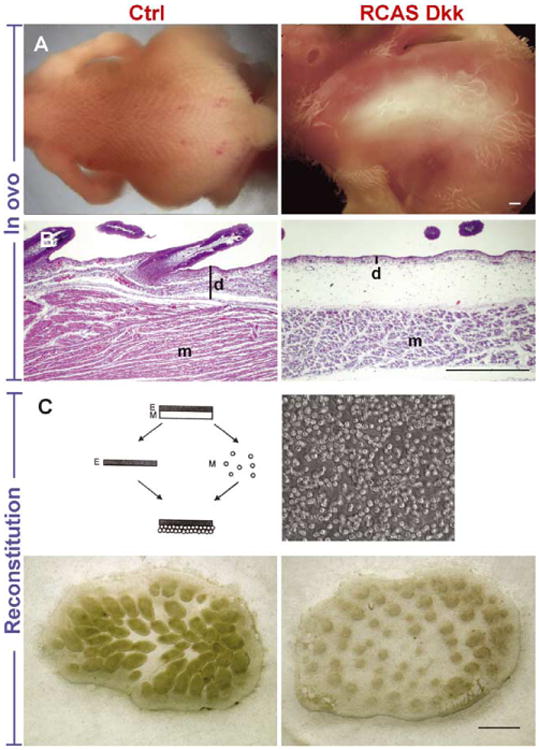
RCAS-DKK inhibited the formation of dermis and feather buds. RCAS-DKK transduction inhibited feather bud formation in vivo and in vitro. RCAS-DKK was injected into the body and amniotic cavity of stage 19–20 (E3) embryos. Feather formation was inhibited over much of the body compared to controls (A). Sections show that feather buds normally elongate with a thickened dermis, but the DKK transduced cultures had a thin dermis (B). RCAS-DKK was then applied to cultured skin explants prepared from stage 31 (E7) chicken embryos. After 4 days in culture, control skin explants formed elongated feather buds, but the feather primordia in DKK transduced cultures remained small and did not elongate (C). Size bars = 500 μm.
To test the direct effect of DKK on individual primordia formation, we transduced reconstituted explant cultures. Dissociated mesenchymal cells were transduced with RCAS-DKK and overlayed with intact epithelium. After 4 days, the cultures showed inhibited feather formation with expanded interbud spacing and smaller feather buds that did not elongate compared to controls (100%, n = 6; Fig. 2C).
Since DKK can suppress formation of the dermis, feather tracts, and individual feather buds, Wnt activities must be involved in the morphogenesis of all of these stages. However, DKK does not differentiate between different Wnt members, and specific antagonists for each of the specific Wnt members do not exist yet. Here we over-expressed each Wnt member to assess their specific function.
Wnt 1
In in ovo assays, ectopic RCAS-Wnt 1 expression can be lethal. Survival to stage 36–40 was about 52% (n = 48). Of these, 16% (n = 4) showed shortened spinal tracts at the lumbosacral level and produced underdeveloped caudal structures by stages 36–40 (E10–E14) (Fig. 3A). Due to this defect, femoral tracts on both sides expanded toward the midline to fill in the gap. Over the abnormal spinal tract, feather buds were either completely absent or loosely arranged with significantly delayed development, resulting in a smaller size (Fig. 3A). Sections of the abnormal tract showed that the ectoderm was intact, but the under developed dermis lost the deep dermal layers and had a thin superficial dermis layer containing few cells. Only loose cells distributed in the extracellular matrix remained compared to controls (Fig. 3B). Collagen II was abundant in the dermal condensations and deep dermis in control skins, but was absent from these regions in Wnt 1-misexpressing samples (Fig. 3B). Tenascin also was missing from this region (data not shown). NCAM weakly stained the superficial dermis, but was dramatically decreased in the deep dermis, compared to controls (Fig. 3B). Antibodies against the RCAS retrovirus (p27) detected virus in the epithelium and dermis (Fig. 3B). They are distributed in irregular patches and at comparable levels as Wnt 3a described below.
In reconstituted explants (n = 12) Wnt 1 transduced explants consistently formed bigger feather buds than controls (Fig. 3C). We then tested the effect of dominant negative Wnt 1 (n = 6). These explants showed thinner feather buds with a similar bud density as controls (Fig. 3C), in contrast to the results of Wnt 1 over-expression. Thus, Wnt 1 misexpression interrupted tract formation by impairing dermis development, which generated smaller or no feather primordia; Wnt 1 induced bigger feather buds on cultured explants.
Wnt 3a
In the in ovo assay, embryos were assessed at stages 36 to 40 (E10–E14). Survival to this stage was low (12/48, 25%), and about 42% of these (n = 5) showed altered feather phenotypes. Affected embryos showed an expanded tract size (there were always 1-2 additional feather bud rows in the dorsal tract on the transduced side compared to the control side) and increased individual feather bud diameter particularly in the caudal tract. To help see the primordia, whole mount in situ hybridization with a Shh probe was used (Fig. 4A). The spinal tracts of RCAS-Wnt 3a transduced specimens were extended into the caudal region, and the dermis was thicker and denser compared to RCAS transduced controls (n = 13). Individual bud size increased as the interbud spacing decreased. Some adjacent buds fused (Fig. 4A). These ‘giant’ buds show expanded distal ends with increased thickness and cellularity in the bud mesenchyme and interbud dermis (Fig. 4B). The virus was distributed in irregular patches (Fig. 4B), but the effect of Wnt 3a may be more widespread due to its non-autonomous function. PCNA staining showed that proliferation was widely dispersed throughout the bud (Fig. 4B).
Fig. 4.
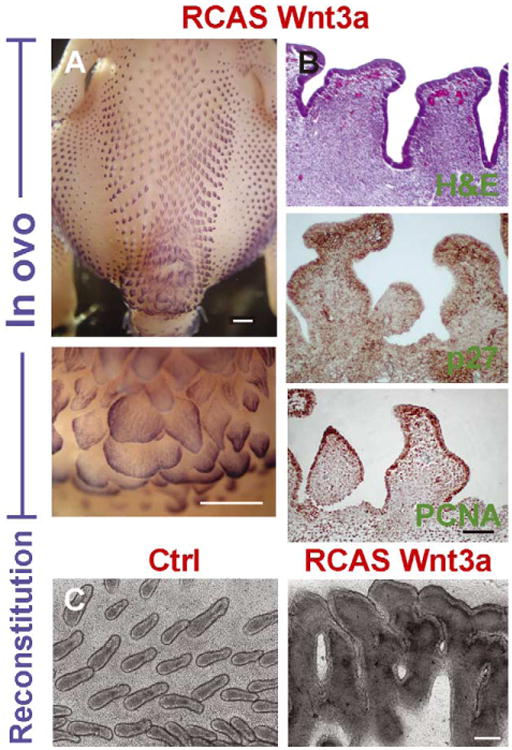
Wnt 3a promotes the formation of feather tracts, expands feather bud size and reduces interbud spacing. RCAS-Wnt 3a transduction induced feather tract expansion, feather primordia fusion or enlargement. Stage 18–22 chicken embryos transduced with RCAS-Wnt 3a virus (right side) in ovo were in situ hybridized to Shh to visualize the feather buds at stage 36–40. The transduced spinal tract was thickened and widened with increased bud size and decreased interbud space (left side) compared to controls (A). An enlarged view of the caudal tract shows that feather primordia were enlarged or fused with neighboring buds at the expense of interbud space. H and E staining showed that RCAS-Wnt 3a infected feather buds had a plateau-like shape with a thickened dermis and increased cellularity (B). RCAS antibody (P27) staining identified the retrovirus in both the epithelium and mesenchyme (B, brown). Diffuse staining of PCNA (B, brown), indicates a wide presence of cell proliferation. RCAS-Wnt 3a transduced reconstituted skin explants had enlarged and fused buds compared to controls (C). Size bars = 500 μm, Size bars for sections = 150 μm.
In reconstituted explants, Wnt 3a increased bud size and bud fusion at the expense of interbud space (n = 4), compared to RCAS controls (n = 4, Fig. 4C). Therefore Wnt 3a misexpression increased dermal thickness, tract size, and individual bud size both in vivo and in vitro.
Wnt 11
In in ovo assays, stage 29 embryos injected with RCAS-Wnt 11 survived to stage 36–40 (E10–14, n = 48). Tract size was not obviously altered when assessed at stage 31–40 (n = 43), but there were focally sparse feather formations and thinning of feather filaments compared to controls (Fig. 5A).
Fig. 5.
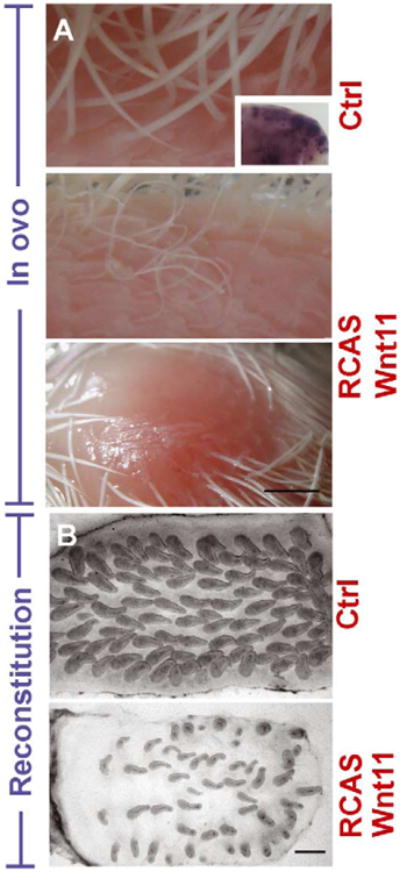
Wnt 11 expands interbud spacing and decreases the feather number and filament diameter. RCAS-Wnt 11 was injected into the body and amniotic cavity of stage 19-20 (E3) embryos. Thinner and sparser feather filaments or focal loss of feathers were detected by stage 40 (E14) in the infected samples compared to controls (A). Control RCAS-alkaline phosphatase transduced embryos stained in wholemount for alkaline phosphatase. RCAS-Wnt 11 infected in vitro reconstituted explants prepared from stage 31 (E7) dorsal skin showed decreased bud size and number, and increased interbud spacing, compared to RCAS-LacZ infected controls (B) after 5 days in culture. Size bars = 500 μm.
In reconstituted skin explants, RCAS-Wnt 11 transduced explants showed decreased number and size of buds and increased interbud spacing, compared with RCAS-LacZ transduced controls (Fig. 5B). Thus Wnt 11 misexpression decreased dermal thickness as well as the number and size of feather buds with increased interbud spacing.
Wnt 5a
RCAS-Wnt 5a also was transduced but did not show apparent results (n = 27). Wnt 5a has been shown to have affects on other studied systems, such as delaying bone maturation in the limb (Hartmann and Tabin, 2000). Its lack of function in the skin is a matter for further investigation.
2.3. Potential relationship among Wnts and different downstream pathways
Next we examined the relationship among Wnt members and searched for their potential downstream pathways. Interesting observations are shown here.
2.3.1. Wnt 3a effects on Wnt 11
We examined the effects of ectopic Wnt 3a expression on the expression of Wnt 11 transcripts (Fig. 6A). In the 5 day control reconstituted explants, Wnt 11 can be seen to be expressed in the interbud mesenchyme (blank arrow) and distal bud mesenchyme (arrow; n = 3). When RCAS-Wnt 3a was transduced in the reconstituted explants, Wnt 3a suppressed Wnt 11 in the interbud and distal bud mesenchyme (Fig. 6A; n = 3). These results are consistent with the notion that Wnt 11 in the interbud is critical for the formation of interbud spacing, while Wnt 11 in the distal bud is important for the tapering of the distal end of elongated feather buds.
Fig. 6.
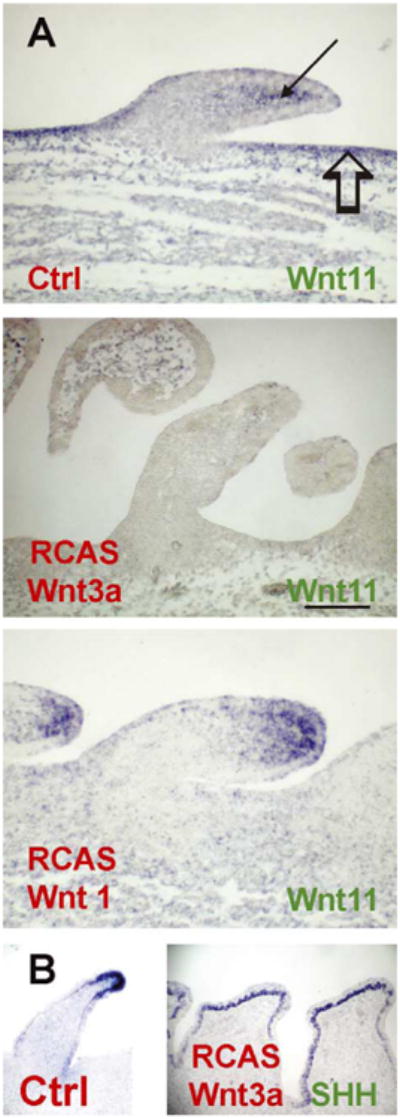
Relationship among molecular pathways. Reconstituted skin explant cultures. (A) At 5 days, endogenous Wnt 11 is expressed in the interbud epithelium (arrow) and distal bud mesenchyme (open arrow). Ectopic Wnt 3a suppresses the expression of Wnt 11 in the interbud region and in the distal bud mesenchyme. In contrast, Wnt 1 induces Wnt 11 in the bud epithelium and mesenchyme but suppresses Wnt 11 in the interbud epithelium. Wnt 3a expands the SHH expressing distal bud growth zone compared to controls (B). Size bar = 100 μm.
2.3.2. Wnt 1 effects on Wnt11
We next examined the effects of ectopic Wnt 1 on the expression of Wnt 11 transcripts (Fig. 6A). In contrast to Wnt 3a, ectopic Wnt 1 induced additional Wnt 11 expression in the distal bud mesenchyme and epithelium (n = 3). It also suppressed normal expression in the interbud epithelium. We do not yet know the mechanism for this differential response, but this may account for the phenotypic differences induced by these two Wnts.
2.3.3. Wnt 3a effects on SHH
We have proposed that the growth of feather buds is driven by a localized growth zone (Chuong et al., 2000). In elongating feather buds, the growth zone is in the distal bud tip and expresses SHH (Ting-Berreth and Chuong, 1996; Fig. 6B). In Wnt 3a transduced feather buds, SHH expression was expanded over the broader distal region and the buds became plateau shaped rather then tapered (Fig. 6B; n = 3).
Since in other systems Wnts signal through several downstream pathways to effect cell behavior, we tested which pathways were used in our system. Although some of the Wnts were expressed by the epithelium, significant effects were observed in the mesenchyme, suggesting that they may signal to targets in the mesenchyme. To test potential mechanisms for these Wnt activities, we tested their roles in cell proliferation, transcriptional activation, PKC activity, cell migration and JNK activity using transduced CEFs in vitro.
2.3.4. Cell proliferation
This was measured as changes in cell number (DNA concentration) over time in control and transduced CEFs (n ≥ 5). Wnt 1, Wnt 3a and Wnt 5a stimulated cell proliferation, while Wnt 11 inhibited cell proliferation compared with controls (Fig. 7A).
Fig. 7.
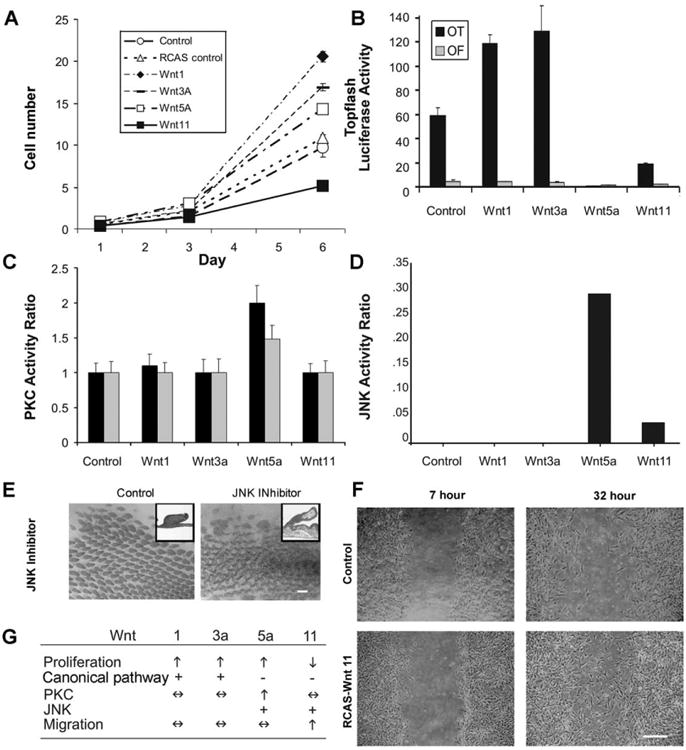
Effect of Wnts on cell proliferation, signaling and migration. (A) Wnt 1, 3a and 5a increased while Wnt 11 inhibited cell proliferation compared to untransduced and RCAS controls. (B) Wnt 1 and Wnt 3a enhanced while Wnt 5a and Wnt 11 suppressed TCF4 promoter activity (OT). No activity was detected with a mutated TCF4 promoter (OF). (C) PKC activity was measured 2 and 4 days after chicken embryo fibroblasts were transduced with retrovirus directing Wnt expression. Only Wnt 5a induced increased PKC activity at these times. The data are presented as a PKC ratio comparing experimental groups to controls at each time point. (D) Wnt 5a and Wnt 11 induced increased JNK activity that was suppressed by SP600125. Three independent experiments were carried out with similar trends. One example is shown. (E) Effect of SP600125, the JNK inhibitor, on skin explant morphogenesis. Control and JNK inhibitor treated skin explants are shown 4 days after treatment. H and E stained cross sections (insets) show that JNK inhibition causes a broadening at the tip of the feather buds. (F) 32 h after scraping mitotically arrested cells from a center stripe, RCAS-Wnt 11 transduced cells reentered the available space faster than control or experimental cell types, demonstrating an increased migration rate. (G) Summary of Wnt effects on skin morphogenesis, molecular pathways and cell behavior. +, activity was detected; −, activity was not detected; ↑, activity was increased; ↓, activity was decreased; ↔, no change in activity. Size bar: E = 50 μm, F = 100 μm.
2.3.5. Beta-catenin/TCF signaling pathway
In the absence of exogenous beta-catenin, cells containing a beta-catenin promoter-luciferase reporter construct had basal levels of luciferase activity. Luciferase activity was significantly increased in RCAS-Wnt 1 and RCAS-Wnt 3a transfected CEFs demonstrating that they signal through the beta-catenin pathway (n ≥ 8). Activation was only observed in the presence of the wildtype binding sequence. RCAS-Wnt 5a and RCAS-Wnt 11 transfected cells had decreased luciferase activity (Fig. 7B), suggesting that they do not signal through the beta-catenin/Tcf pathway. Their ectopic expression even suppressed the canonical pathway in CEFs.
2.3.6. PKC activity
RCAS-Wnt 5a transfected cells cultured for 2 and 4 days showed consistent activation of PKC activity (Fig. 7C, n ≥ 8). Therefore, Wnt 5a can activate the PKC pathway in CEFs but Wnt 1, Wnt 3a, and Wnt 11 did not.
2.3.7. JNK activity
Cell extracts were electrophoresed and then immunoblotted with an anti-JNK 1/2 polyclonal antibody (Biomol). The intensity of bands at the appropriate molecular weight as assessed with a BioRad Fluor-S multimager, that decreased in cultures grown in the presence of SP600125, a specific JNK-1, -2, and -3 inhibitor were considered to be representative of JNK activity (n ≥ 6). Wnt 5a induced the most kinase activity suppressible by the JNK inhibitor. Wnt 11 induced kinase activity with the correct size on a western blot that was partially suppressed by the JNK inhibitor (Fig. 7D).
We next tested the effects of the JNK inhibitor, SP600125, on cultured skin explants. E8 dorsal skin was cultured in the presence or absence of 10 μM SP600125 for 4 days (Fig. 7E). The bud regions (especially the peripheral bud regions) expanded at the expense of the interbud regions (n = 4; data not shown). This is particularly obvious for feather primordia toward the lateral edge of the explant that had not initiated their formation when inhibitors were added. Feather buds near the midline which had finished their periodic patterning when inhibitors were added showed enlarged distal feather buds, rather than the tapering bud tips present in controls. These data suggest that JNK is involved in establishing interbud spacing and for keeping the distal feather bud tips tapered. This JNK activity may be induced by Wnt 5a, 11 or others.
2.3.8. Cell migration
To test the effect of cell migration in cultured cells, the proliferation of confluent transduced cells first was inhibited by mitomycin C, then a line of cells was scraped from the dish (n = 3). Tracking cell migration back into the scraped region at 0, 7, 24, and 32 h showed that Wnt 11 consistently and significantly stimulated cell migration (Fig. 7F), while the other Wnts did not compared to controls (data not shown). The results are summarized in Fig. 7G.
3. Discussion
The current study demonstrates that different Wnt members play positive or negative roles at three successive levels of skin morphogenesis: during the formation of the (1) skin regions (e.g. feather tracts), (2) individual primordia, and (3) individual appendage shapes (Fig. 8). Wnts were previously shown to exert an affect on dermis formation (Olivera-Martinez et al., 2001, 2002). Skin morphogenesis occurs at different hierarchical levels (Chuong et al., 2000; Harris et al., 2002; Prum and Dyck, 2003). These analyses help us understand why the same molecule can elicit different results when altered at different developmental times. For instance, when Shh was misexpressed early it induced disorganized epithelial proliferation, whereas it later induced feather bud formation and still later played a role in feather branch apoptosis (Morgan et al., 1998; Ting-Berreth and Chuong, 1996; Harris et al., 2002; Yu et al., 2002). Stage specific differences were also seen with BMP 2, which suppressed individual feather primordia formation (Jung et al., 1998; Noramly and Morgan, 1998), but also induced an ectopic feather tract when applied at different stages (Scaal et al., 2002). While analysis at different levels helps to clarify the morphogenetic process, in actuality, each developmental process progresses directly into the next without a clear pause. Therefore the different experimental designs used here are interpreted as having major effects on certain levels of morphogenesis, but they probably influence other levels, as well.
Fig. 8.
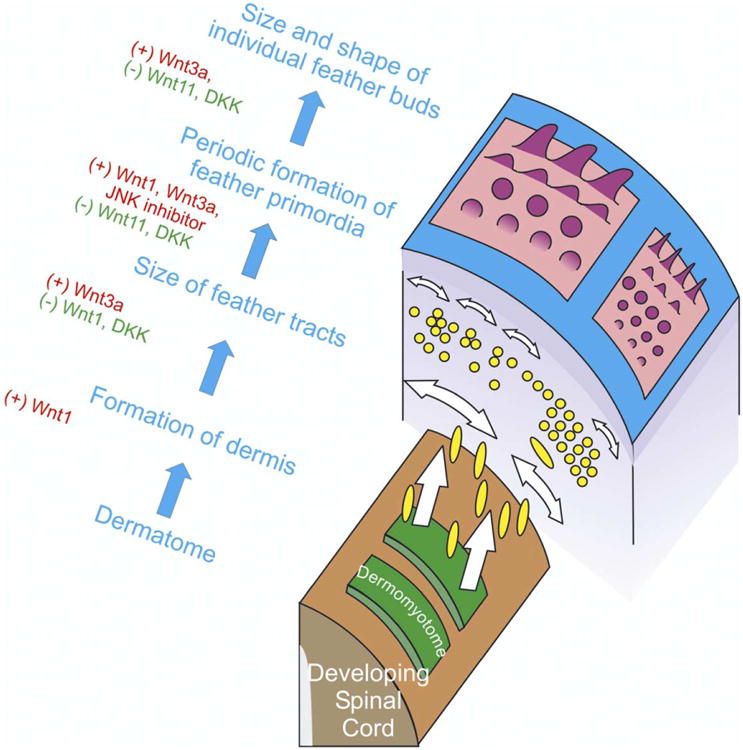
Schematic summary of the positive and negative roles of Wnts during different hierarchical levels of skin morphogenesis. Four morphogenetic levels between five structural entities (dermomyotome, dermis, feather tracts, individual feather primordia, characteristic feather shape) are shown. Yellow spindles and circles represent dermal cells. Blue represents the epidermis. Two different tracts with different density and shape of skin appendages are shown. At each morphogenetic level, Wnts play either a positive (red) or negative role (green). Data for the dermatome to dermis migration is based on Olivera-Martinez et al., 2001, 2002.
3.1. Wnts regulate diverse signaling pathways in dermal fibroblasts
The formation of dermis, tract and individual primordia are regulated by cell migration, proliferation, and adhesion of dermal precursor cells. Wnts activate a number of downstream pathways influencing these behaviors in other experimental systems (Wang and Malbon, 2003). In fact, the same Wnt ligands may activate several independent signaling pathways, including beta-catenin dependent gene expression, morphogenetic movements, or intracellular calcium release. These diverse cell responses may depend on the availability of specific frizzled receptors or coreceptors, differences in binding affinity between ligands and receptors, or on the concentration of downstream pathway components.
While Wnt 1 and Wnt 3a were expressed in the epithelium, their predominant effects are seen in the dermis. Since Wnts are known to act as paracrine signals, this result is not surprising. Therefore, we tested their function using chicken embryo fibroblasts. The cellular responses to Wnt stimulation are summarized (Fig. 7G). Wnt 1 and Wnt 3a stimulated cell proliferation by triggering the canonical beta-catenin pathway. This suggests that they may be playing similar roles in vivo, but we cannot rule out the possibility that they also signal through non-canonical pathways. The specific frizzled receptors utilized by the different Wnts have not been ascertained as yet. Noramly et al. (1999) report that beta-catenin is transiently expressed in the nucleus of dense dermis at early stages of feather bud initiation. Wnt 11 stimulated cell migration and inhibited cell proliferation, possibly by blocking the beta-catenin pathway. These results are consistent with the notion that Wnt 1 and Wnt 3a favor the formation of a feather primordium domain, while Wnt 11 suppresses feather bud formation.
Wnt 5a and Wnt 11 exerted their effects through non-canonical Wnt pathways by triggering the JNK pathway. Wnt 5a also increased PKC activity. This suggests that they both signal throught non-canonical pathways in vivo, but we cannot rule out the possibility that they also can induce the canonical pathway in vivo. Both Wnt 5a and Wnt 11 were initially expressed in the interbud epithelium and dermis, respectively and later in the bud mesenchyme. We propose that their interbud expression helps establish interbud spacing, while the distal bud expression keeps the growing bud tips tapered. The finding that in feather explant cultures the JNK inhibitor decreased the size of the interbud spacing in the lateral (younger) regions, but expanded the size of distal buds in the center (more mature) is consistent with this explanation. Increased cell migration induced by Wnt 11 is antagonistic to dermal condensation formation, which requires higher cell adhesion (Jiang and Chuong, 1992) or accumulation of cells in the bud tips. It is not clear why Wnt 5a failed to produce a phenotypic change. Wnt 5a moderately stimulated cell proliferation, triggered the PKC and JNK pathways and inhibited the beta-catenin pathway. This is consistent with its functions in other systems, where it increased the proliferation of progenitor cells in mouse development (Yamaguchi et al., 1999), triggered intracellular Ca+2 release to activate PKC and CamKII (Sheldahl et al., 1999; Kühl et al., 2000), decreased cell adhesion (Torres et al., 1996) and inhibited the Wnt/beta-catenin pathway (Kühl et al., 2001; Ishitani et al., 2003).
3.2. Wnt and the formation of dermis
In avian embryos the dermis at different locations within the body are derived from different origins. The ventral and lateral trunk dermis and limb bud mesenchyme are derived from the somatopleura and the dorsal dermis is from the dermomyotome of the somite (Mauger, 1972). Dermis in the head comes from the cephalic crest, paraxial mesoderm, lateral plate mesoderm and prechordal mesoderm (Noden, 1991). Here we discuss the dermatome-dorsal tract dermis.
Wnt 1 and Wnt 3a secreted from the dorsal neural tube induce dermomyotome specification and/or maintenance (Capdevila et al., 1998; Ikeya and Takada, 1998; Wagner et al., 2000). Replacing the neural tube with cells expressing Wnt 1, but not Wnt 3a, BMP2 nor noggin, produced a feather forming dermis, demonstrating that Wnt 1 alone allows the survival and specification of the medial dermomyotome compartment (Olivera-Martinez et al., 2001). Wnt 1 up-regulates Wnt 11 in the dermomyotome (Capdevila et al., 1998). Wnt 11 expressing cells in the dorsomedial lip of the dermomyotome migrate under the ectoderm, towards the midline, to form most of the dermis (Olivera-Martinez et al., 2002). In parallel, we found that ectopic Wnt 1 expression increased Wnt 11 expression in transduced embryos and that Wnt 11 increased cell migration in CEFs. Therefore, Wnt 1 may promote dermal precursor cells to migrate from the dermatome to the dermis by inducing Wnt 11.
3.3. Wnt and the formation of skin regions (macropatterning)
During dermis formation, presumptive dermal cells in the subectoderm sort out to form different skin regions. These skin regions are clearly demarcated in birds, as feather tracts distributed over the body surface, separated by apteric regions (Lucas and Stettenheim, 1972) in a process called macropatterning (Sengel, 1976).
Wnt 1 and Wnt 3a normally were expressed throughout the tract epidermis but not in the apteric regions. Wnt 11, on the other hand, is in both the tract and apteria, transiently (Tanda et al., 1995). In our experiments genes are misexpressed globally by retroviral vector injection to the amniotic cavity at stage 12–20 (E2/E3). Wnt 1 and Wnt 3a are thought to function through similar canonical beta-catenin pathways and yet mis-expression to similar levels exerted opposite effects on both tract size and dermal thickness. Wnt 1 produced a truncated spinal tract and the neighboring femoral tracts expanded into the affected sacral-caudal region. On the other hand, Wnt 3a induced a caudally expanded spinal tract. Sections of Wnt 3a misexpressing samples had a thickened dermis, higher cellularity, and an increase in the PCNA-positive cell number (data not shown) indicating active proliferation. Wnt 1 over expressing skin did not express collagen II or NCAM. Sections revealed a thin layer of superficial dermis with decreased or absent dermal condensations. The disparate effects of Wnt 1 and Wnt 3a on tract formation may be attributed to their differential regulation of downstream molecular expression and subsequent cell behavior, although it is unknown if these effects are mediated through distinct frizzled receptors leading to activation of the canonical or non-canonical pathways. While the disparate effects of Wnt 1 and Wnt 3a are the subject of future studies, Wnt 3a suppresses Wnt 11, while Wnt 1 induces Wnt 11, which in turn promotes cell migration and may partially inhibit cell proliferation. Perhaps Wnt 3a enhances tract formation by increasing cell numbers in tract regions via cell proliferation induction and cell migration suppression. Wnt 1 may suppress early tract formation through Wnt 11 mediated maintenance of migration leading to suppression of proliferation and differentiation in dermal cells. However, Wnt 11 alone, examined at stages 31–40 (data not shown), had no apparent effect on tract formation at this stage, suggesting that it is functioning in conjunction with some other induced gene product. Hence, Wnt functions appear to be mediated through the combined activities of several downstream pathways. When frizzled mediated Wnt signaling was inhibited with DKK 1, the dermis remained thin. This suggests that Wnt signaling is required for dermis formation.
3.4. Wnt and the induction of individual feather primordia (micropatterning)
Feather primordia appear in sequence and through micropatterning are arranged in distinct patterns within feather tracts (Sengel, 1976; Mayerson and Fallon, 1984). Molecules regulating this process are expressed in two modes. In the restrictive mode, molecules appear early, initiating throughout the tract and then gradually become restricted to the bud or interbud. These molecules are involved in sorting out bud and interbud domains. In the de novo mode, molecules appear later, directly in the bud or interbud and are involved in consolidating the bud/interbud domains and subsequent bud morphogenesis. All of the Wnts examined here are expressed in a restrictive mode. They become restricted in three general expression patterns. Following the first pattern, Wnt 1 and 3a are initially expressed throughout the epithelium and later become restricted to the bud epithelium with a similar time course. Following the second pattern, Wnt 11 is always in the mesenchyme and becomes restricted to the interbud region, and, to a minor extent, the distal bud. Following the third pattern, Wnt 5a is in the epithelium and switches to the mesenchyme. Later it is in the distal bud mesenchyme. These different expression patterns suggest at least three distinct roles. However, while Wnt 1 and 3a show some functional similarities, they also have disparate affects. Their shared expression patterns suggest that they either may have overlapping functions or modulate each other's functions.
Perturbation of molecular expression can alter the size and spacing of the early primordia (Jiang et al., 1999). Molecules can be generally categorized as activators or inhibitors for bud formation. Through mechanisms involving a reaction–diffusion mechanism, regions with a higher activator to inhibitor ratio build a bud rather than an interbud, and vice versa (Jiang et al., 1999). Using a dermal cell reconstitution assay which resets epithelial and mesenchymal cells to an early step in dermis formation (Jiang et al., 1999), we showed that Wnt 3a is an activator, generating big bud domains that sometimes fused into stripes. Wnt 1 also was an activator promoting large but discrete buds, while a dominant negative Wnt 1 produced thinner feather buds. The activator function is consistent with activation of the canonical beta-catenin pathway. However, the activator function contrasts with its effect in ovo. The reconstitution assay used here was performed on a normal stage 31 (E7) dermis and focuses on factors effecting feather primordia induction rather than dermis formation. This suggests that at this stage, the main effect of Wnt 1 and Wnt 3a expression are exerted through the canonical beta-catenin pathway. Wnt 11 is an inhibitor generating fewer and smaller feather buds in this assay. This means that at this step, Wnt 11 can function in the absence of the molecular partner(s) that are required to affect skin region formation. Over-expression of Wnt 11 may increase cell migration and inhibit condensation of dermal cells under individual feather primordia. It is possible that Wnt 11 expressing cells then become interbud and Wnt 11 non-expressing cells become bud, although causality requires further study.
3.5. Wnt and the shape of feather buds
As skin appendages grow, they assume different shapes. We proposed that localized cell proliferation and cell death may mediate these shaping processes to form specific designs for special functions (Chuong et al., 2000). Here we propose that the shape of feather buds is regulated by the size of this distal bud growth zone that is SHH positive. Wnt 3a works as a positive regulator to expand this growth zone, presumably through the canonical beta-catenin pathway. JNK activity, through Wnt 5a, 11, or others works as a negative regulator to make this zone small. A normal balance leads to a feather bud shape that tapers toward the distal end. When positive regulators dominate, the bud tips remain the same diameter as the base and become ‘giant buds’ with an expanded distal end. When negative regulators dominate, the buds taper too fast, and the diameter of feather filaments become very thin.
In normal conditions, as the buds begin to elongate, the growth zone disappears from the distal tip and starts to shift down to the proximal end (Chuong et al., 2000). The persistent existence of a growth zone in the distal end leads to a diffuse distribution of proliferating cells which does not promote continuous bud elongation. This can be seen in Wnt 3a buds, and also in the Wnt7a over-expressing buds (Widelitz et al., 1999). The shift of the growth zone toward the proximal feather follicle is regulated by Wnt signaling (e.g. Wnt 6, Wnt 14, Sfrp 2). Localized misexpression of Wnt 6 causes a localized enlargement of feather filaments (Chodankar et al., 2003). There are more hierarchical levels of morphogenesis that eventually establish the complex branching patterns of feathers (Prum and Dyck, 2003; Yu et al., 2002). Wnt pathway members (e.g. Wnt 6, 8c, Sfrp 2) are also expressed in feather filaments and may be involved in these processes (Chodankar et al., 2003).
Our studies showed that chicken skin morphogenesis can be better understood if analyzed in hierarchical levels: from the dermatome to subectodermal dermis, to skin regions, to individual skin appendages, and finally to characteristic appendage design (Fig. 8). At each level, Wnts or other signaling molecules may have different effects by altering cell behaviors (migration, adhesion, proliferation, differentiation, etc).
4. Experimental procedures
4.1. Recombinant retroviral construction and production
SPF-chick embryo fibroblast (CEF) cultures were grown and transfected with RCAS-Wnt 1, -Wnt 3a, -Wnt 5a, -Wnt 11 and -DKK 1 retroviral vector DNAs or control RCAS vector as described by Morgan and Fekete (1996). RCAS Wnt 1, 3a was from Dr Tabin's lab (Kengaku et al., 1998). RCAS Wnt 5a, 11 were from Dr Nohno's lab (Kawakami et al., 1999; Anakwe et al., 2003). Mouse DKK 1 in pGEM-T Easy from Drs Lassar and Millar (Andl et al., 2002) was excised with a partial NcoI, EcoRI digestion and subcloned into the pENTR 1A Gateway entry vector (Invitrogen). A recombinase reaction was used to transfer the mouse DKK 1 cDNA to an RCAS vector prepared for the Gateway cloning system (Loftus et al., 2001; Invitrogen). The supernatants of CEF cultures transfected with the viral DNAs were aliquoted and stored at −80 °C until use. RCAS-DKK (Dickkopf) was constructed as described in Suksaweang et al., (2004).
4.2. RCAS in ovo injection
RCAS viruses were injected in ovo to the amniotic cavity or embryonic trunk at stage 12–20 (E2 or E3). The presence of virus was verified by staining for the retroviral gag gene, capsid protein or by staining for the transduced gene product.
4.3. Skin reconstitution assay in vitro
Dissociated mesenchymal cells from stage 30 chicken embryos were resuspended in DMEM and plated at high cell density, then covered with an intact piece of stage 30 epithelium to form reconstituted explants (Fig. 2; Jiang et al., 1999). The explants were cultured on tissue culture inserts (Falcon) for the designated number of days. For gene misexpression, dissociated mesenchyme cells were resuspended in virus containing media. RCAS-LacZ transduced or non-viral reconstituted feather explants were used as controls. For dominant negative Wnt 1 (DN Wnt 1) experiments, DN Wnt 1 producing cell lines were used (Garcia-Castro et al., 2002). Reconstituted explants were co-cultured on culture inserts with DN Wnt 1 producing cells grown on the bottom of the dish. 3T3 cells and conditioned media were used as controls.
4.4. In situ hybridization and immunohistochemistry
Specific pathogen-free (SPF) White Leghorn chick embryos were obtained from SPAFAS (Preston, CT). Embryos staged according to Hamburger and Hamilton (HandH) (1951) were fixed in 4% paraformaldehyde and processed for RNA whole mount in situ hybridization, section in situ hybridization or immunohistochemistry (Ting-Berreth and Chuong, 1996; Jiang and Chuong, 1992). Antibodies against RCAS p27 (SPAFAS), collagen II (Sigma), and a polyclonal antibody to NCAM were used for immunohistochemistry.
4.5. Proliferation assay
Cells were cultured in 60 mm dishes with three replicates per experiment The DNA concentration was measured on Day 1, 3, and 6. Cells were washed twice with cold calcium-magnesium-free phosphate-buffered saline (CMF-PBS), extracted twice with cold methanol: acetic acid (3:1) to remove nucleosides and nucleotides, and treated at 37 °C with 0.2N NaOH at 1 × 106 cells per ml for 24 h. The absorption at 260 nm for each cell solution was determined with a Bio-Rad 3000 spectrophotometer. Each point represents the average of five independent experiments.
4.6. Luciferase assay for TCF
Transfection was done according to the manufacturer's instructions (Targeting systems). CEFs were cultured in 12-well dishes to approximately 70% confluence and washed two times with OptiMem before adding transfection mixes. Transfection mix containing OptiMem, plasmid DNA and Targefect F2 was incubated at 37 °C for 20 min to form a transfection complex. This was added to each well, incubated at 37 °C for 2 h before adding serum-containing DMEM. Luciferase activity was assayed 24 h after transfection following the manufacturer's instructions (Promega). Two different TCF-luciferase reporter constructs were used: a wildtype TCF-luciferase reporter construct (pGL3-OT) and a mutated TCF-luciferase reporter construct (pGL3-OF), which served as a negative control (He et al., 1998). Each bar graph shows the average of four independent experiments.
4.7. Cell migration assay
When cells reached 100% confluence, 10 μg/ml mitomycin C (Sigma) was added and incubated for 3-hour to inhibit proliferation. Then a line of cells was scraped from the culture dish. Cell migration back into the scraped zone was recorded at 0, 7, 24, and 32 h by digital photography using a stereoscopic microscope (Nikon). Each condition was repeated in triplicate.
4.8. Protein kinase assay
PKC acivity was measured according to the manufacturer's instructions (Protein Kinase Assay Kit, Calbiochem). CEFs treated with 1 μm PMA, or 100 nm calphostin C were used for positive and negative controls, respectively. The assay kit was based on an ELISA, utilizing a synthetic PKC pseudosubstrate and a monoclonal antibody that recognizes the phosphorylated form of the peptide. The OD at 450 nm was determined in a microplate reader. Each point represents the average of four independent experiments.
4.9. JNK assay
CEFs were treated for 24 h with 0.1% DMSO or 10 μM SP600125 (prepared in 0.1% DMSO), a potent and selective JNK-1, -2, -3 inhibitor that suppresses the phosphorylation of c-jun (Biomol). CEF cell extracts were separated by 10% SDS-polyacrylamide gel electrophoresis, transferred to nitrocellulose and immunoblotted with anti-JNK 1/2 (pTpY 183/185), a rabbit polyclonal antibody (Biomol). JNK activity was assessed as an immunoprecipitated band whose appearance was suppressed by SP600125. The intensity of the immunoprecipitated band was assessed using a BioRad Fluor-S multimager. This experiment was performed three times. The data presented shows one representative sample.
Dorsal chicken skin explants were prepared from stage 34 embryos and exposed to SP600125 (10 μM) for 4 days. Skin explants exposed to the JNK inhibitor were photographed daily to document alterations of feather formation. Controls were exposed to 0.1% DMSO.
Acknowledgments
This work was supported by grants from the US NIH (RW, CMC, LWB), US National Science Foundation (CMC), National Science Council in Taiwan (CHC), and Tzu Chi Medical Center in Taiwan (CHC). We thank the following for providing various reagents. In situ probes and RCAS retroviral vectors were from Dr McMahon (Harvard University, Cambridge, MA), Dr Nohno (Kawasaki Medical School, Okayama, Japan) and Dr Tabin (Harvard University, Cambridge, MA). DN Wnt 1 producing cells were kindly provided by Dr Garcia-Castro and Dr Bronner-Fraser (California Institute of Technology). DKK was from Dr Andrew Lassar (Harvard University, Cambridge, MA) and Dr Sarah Millar (University of Pennsylvania, Philadelphia, PA). The pGL3-OT and pGL3-OF TCF4 reporter constructs were from Dr Bert Vogelstein (Johns Hopkins University, Baltimore, MD). We thank Mr Michael W. Hughes for making RCAS-DKK 1 construct. We thank Dr Hsin-Su Yu for helpful discussion during the preparation of the manuscript.
References
- Anakwe K, Robson L, Hadley J, Buxton P, Church V, Allen S, et al. Wnt signalling regulates myogenic differentiation in the developing avian wing. Development. 2003;130:3503–3514. doi: 10.1242/dev.00538. [DOI] [PubMed] [Google Scholar]
- Andl T, Reddy ST, Gaddapara T, Millar SE. WNT signals are required for the initiation of hair follicle development. Dev Cell. 2002;2:643–653. doi: 10.1016/s1534-5807(02)00167-3. [DOI] [PubMed] [Google Scholar]
- Capdevila J, Tabin C, Johnson R. Control of dorsoventral somite patterning by Wnt 1 and beta-catenin. Dev Biol. 1998;193:182–194. doi: 10.1006/dbio.1997.8806. [DOI] [PubMed] [Google Scholar]
- Chen CW, Chuong CM. Avian integument provides multiple possibilities to analyse different phases of skin appendage morphogenesis. J Investig Dermatol Symp Proc. 1999;4:333–337. doi: 10.1038/sj.jidsp.5640240. [DOI] [PubMed] [Google Scholar]
- Chodankar R, Chang CH, Yue Z, Jiang TX, Suksaweang S, Burrus LW, et al. Shift of localized growth zones contributes to skin appendage morphogenesis: role of the Wnt/beta-catenin pathway. J Investig Dermatol. 2003;120:20–26. doi: 10.1046/j.1523-1747.2003.12008.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuong CM. The making of a feather: homeoproteins, retinoids and adhesion molecules. Bioessays. 1993;15:513–521. doi: 10.1002/bies.950150804. [DOI] [PubMed] [Google Scholar]
- Chuong CM, Edelman GM. Expression of cell-adhesion molecules in embryonic induction, I. Morphogenesis of nestling feathers. J Cell Biol. 1985;101:1009–1026. doi: 10.1083/jcb.101.3.1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuong CM, Widelitz RB. Feather morphogenesis: a model of the formation of epithelial appendages. In: Chuong CM, editor. Molecular Basis of Epithelial Appendage Morphogenesis. Landes Bioscience; Austin: 1998. pp. 57–74. [Google Scholar]
- Chuong CM, Ting-Berreth S, Widelitz RB, Jiang TX. Early events during the regeneration of skin appendages: order of molecular reappearance following epithelial–mesenchymal recombination with rotation. J Invest Dermatol. 1996;107:639–646. doi: 10.1111/1523-1747.ep12584254. [DOI] [PubMed] [Google Scholar]
- Chuong CM, Chodankar R, Widelitz RB, Jiang TX. Evo–Devo of feathers and scales: building complex epithelial appendages. Curr Opin Genet Dev. 2000;10:449–456. doi: 10.1016/s0959-437x(00)00111-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhouailly D. Feather-forming capacities of the avian extra-embryonic somatopleure. J Embryol Exp Morphol. 1978;43:279–287. [PubMed] [Google Scholar]
- Dierick H, Bejsovec A. Cellular mechanism of wingless/Wnt signal transduction. Curr Top Dev Biol. 1999;43:153–190. doi: 10.1016/s0070-2153(08)60381-6. [DOI] [PubMed] [Google Scholar]
- Garcia-Castro MI, Marcelle C, Bronner-Fraser M. Ectodermal Wnt function as a neural crest inducer. Science. 2002;297:848–851. doi: 10.1126/science.1070824. [DOI] [PubMed] [Google Scholar]
- Hamburger V, Hamilton HL. A series of normal stages in the development of the chick embryo. J Morphol. 1951;88:49–92. [PubMed] [Google Scholar]
- Harris MP, Fallon JF, Prum RO. Shh-Bmp2 signaling module and the evolutionary origin and diversification of feathers. J Exp Zool. 2002;294:160–176. doi: 10.1002/jez.10157. [DOI] [PubMed] [Google Scholar]
- Hartmann C, Tabin CJ. Dual roles of Wnt signaling during chondrogenesis in the chicken limb. Development. 2000;127:3141–3159. doi: 10.1242/dev.127.14.3141. [DOI] [PubMed] [Google Scholar]
- He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, et al. Identification of c-MYC as a target of the APC pathway. Science. 1998;281:1509–1512. doi: 10.1126/science.281.5382.1509. [DOI] [PubMed] [Google Scholar]
- Ikeya M, Takada S. Wnt signaling from the dorsal neural tube is required for the formation of the medial dermomyotome. Development. 1998;125:4969–4976. doi: 10.1242/dev.125.24.4969. [DOI] [PubMed] [Google Scholar]
- Ishitani T, Kishida S, Hyodo-Miura J, Ueno N, Yasuda J, Waterman M, et al. The TAK1-NLK mitogen-activated protein kinase cascade functions in the Wnt-5a/Ca2+ pathway to antagonize Wnt/beta-catenin signaling. Mol Cell Biol. 2003;23:131–139. doi: 10.1128/MCB.23.1.131-139.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang TX, Chuong CM. Mechanism of skin morphogenesis. I. Analysis with antibodies to adhesion molecules tenascin, N-CAM, and integrin. Dev Biol. 1992;150:82–98. doi: 10.1016/0012-1606(92)90009-6. [DOI] [PubMed] [Google Scholar]
- Jiang TX, Jung HS, Widelitz RB, Chuong CM. Self-organization of periodic patterns by dissociated feather mesenchymal cells and regulation of size, number and spacing of primordial. Development. 1999;126:4997–5009. doi: 10.1242/dev.126.22.4997. [DOI] [PubMed] [Google Scholar]
- Jung HS, Francis-West PH, Widelitz RB, Jiang TX, Ting-Berreth S, Tickle C, et al. Local inhibitory action of BMPs and their relationships with activators in feather formation: implications for periodic patterning. Dev Biol. 1998;196:11–23. doi: 10.1006/dbio.1998.8850. [DOI] [PubMed] [Google Scholar]
- Kawakami Y, Wada N, Nishimatsu SI, Ishikawa T, Noji S, Nohno T. Involvement of Wnt-5a in chondrogenic pattern formation in the chick limb bud. Dev Growth Differ. 1999;41:29–40. doi: 10.1046/j.1440-169x.1999.00402.x. [DOI] [PubMed] [Google Scholar]
- Kengaku M, Capdevila J, Rodriguez-Esteban C, De La Pena J, Johnson RL, Belmonte-Ispisua JC, Tabin CJ. Distinct WNT pathways regulating AER formation and dorsoventral polarity in the chick limb bud. Science. 1998;280:1274–1277. doi: 10.1126/science.280.5367.1274. [DOI] [PubMed] [Google Scholar]
- Kratochwil K, Dull M, Farinas I, Galceran J, Grosschedl R. Lef1 expression is activated by BMP-4 and regulates inductive tissue interactions in tooth and hair development. Genes Dev. 1996;10:1382–1394. doi: 10.1101/gad.10.11.1382. [DOI] [PubMed] [Google Scholar]
- Kühl M, Sheldahl LC, Park M, Millar JR, Moon RT. The Wnt/Ca2+ pathway: a new vertebrate Wnt signaling pathway takes shape. Trends Genet. 2000;16:279–283. doi: 10.1016/s0168-9525(00)02028-x. [DOI] [PubMed] [Google Scholar]
- Kühl M, Geis K, Sheldahl LC, Pukrop T, Moon RT, Wedlich D. Antagonistic regulation of convergent extension movements in Xenopus by Wnt/β-catenin and Wnt/Ca2+ signaling. Mech Dev. 2001;106:61–76. doi: 10.1016/s0925-4773(01)00416-6. [DOI] [PubMed] [Google Scholar]
- Loftus SK, Larson DM, Watkins-Chow D, Church DM, Pavan WJ. Generation of RCAS vectors useful for functional genomic analyses. DNA Res. 2001;8:221–226. doi: 10.1093/dnares/8.5.221. [DOI] [PubMed] [Google Scholar]
- Lucas AM, Stettenheim PR. Agriculture handbook, Agricultural research sciences. Vol. 362. US Department of Agriculture; Washington, DC: 1972. Avian anatomy. Integument. [Google Scholar]
- Lustig B, Behrens J. The Wnt signaling pathway and its role in tumor development. J Cancer Res Clin Oncol. 2003;129:199–221. doi: 10.1007/s00432-003-0431-0. [DOI] [PubMed] [Google Scholar]
- Malbon CC, Wang H, Moon RT. Wnt signaling and heterotrimeric G-proteins: strange bedfellows or a classic romance? Biochem Biophys Res Commun. 2001;287:589–593. doi: 10.1006/bbrc.2001.5630. [DOI] [PubMed] [Google Scholar]
- Mauger A. The role of somatic mesoderm in the development of dorsal plumage in chick embryos. I. Origin, regulative capacity and determination of the plumage-forming mesoderm. J Embryol Exp Morphol. 1972;28:313–341. [PubMed] [Google Scholar]
- Mayerson PL, Fallon JF. The effects of Janus Green B on the temporal and spatial pattern of feather germ morphogenesis. J Exp Zool. 1984;232:231–242. doi: 10.1002/jez.1402320210. [DOI] [PubMed] [Google Scholar]
- Millar SE, Willert K, Salinas PC, Roelink H, Nusse R, Sussman DJ, Barsh GS. Wnt signaling in the control of hair growth and structure. Dev Biol. 1999;207:133–149. doi: 10.1006/dbio.1998.9140. [DOI] [PubMed] [Google Scholar]
- Morgan BA, Fekete DM. Manipulating gene expression with replication-competent retroviruses. Methods Cell Biol. 1996;51:185–218. doi: 10.1016/s0091-679x(08)60629-9. [DOI] [PubMed] [Google Scholar]
- Morgan BA, Orkin RW, Noramly S, Perez A. Stage-specific effects of sonic hedgehog expression in the epidermis. Dev Biol. 1998;201:1–12. doi: 10.1006/dbio.1998.8969. [DOI] [PubMed] [Google Scholar]
- Noden DM. Cell movements and control of patterned tissue assembly during craniofacial development. J Craniofac Genet Dev Biol. 1991;11:192–213. [PubMed] [Google Scholar]
- Noramly S, Morgan BA. BMPs mediate lateral inhibition at successive stages in feather tract development. Development. 1998;125:3775–3787. doi: 10.1242/dev.125.19.3775. [DOI] [PubMed] [Google Scholar]
- Noramly S, Freeman A, Morgan BA. beta-catenin signaling can initiate feather bud development. Development. 1999;126:3509–3521. doi: 10.1242/dev.126.16.3509. [DOI] [PubMed] [Google Scholar]
- Olivera-Martinez I, Thélu J, Teillet MA, Dhouailly D. Dorsal dermis development depends on a signal from the dorsal neural tube, which can be substituted by Wnt-1. Mech Dev. 2001;100:233–244. doi: 10.1016/s0925-4773(00)00540-2. [DOI] [PubMed] [Google Scholar]
- Olivera-Martinez I, Missier S, Fraboulet S, Thelu J, Dhouailly D. Differential regulation of the chick dorsal thoracic dermal progenitors from the medial dermomyotome. Development. 2002;129:4763–4772. doi: 10.1242/dev.129.20.4763. [DOI] [PubMed] [Google Scholar]
- Prum RO, Dyck J. A hierarchical model of plumange: morphology, development and evolution. J Exp Zool. 2003;298:73–90. doi: 10.1002/jez.b.27. [DOI] [PubMed] [Google Scholar]
- Scaal M, Pröls F, Füchtbauer EM, Patel K, Hornik C, Köhler T, et al. BMPs induce dermal markers and ectopic feather tracts. Mech Dev. 2002;110:51–60. doi: 10.1016/s0925-4773(01)00552-4. [DOI] [PubMed] [Google Scholar]
- Sengel P. Morphogenesis of Skin. Cambridge University Press; Cambridge: 1976. [Google Scholar]
- Sheldahl LC, Park M, Malbon CC, Moon RT. Protein kinase C is differentially stimulated by Wnt and Frizzled homologs in a G protein-dependent manner. Curr Biol. 1999;9:695–698. doi: 10.1016/s0960-9822(99)80310-8. [DOI] [PubMed] [Google Scholar]
- Suksaweang S, Lin CM, Jiang TX, Hughes MW, Widelitz R, Chuong CM. Morphogenesis of chicken liver: identification of localized growth zones and the role of beta-catenin/Wnt in size regulation. Dev Biol. 2004 doi: 10.1016/j.ydbio.2003.10.010. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanda N, Ohuchi H, Yoshioka H, Noji S, Nohno T. A chicken Wnt gene, Wnt 11, is involved in dermal development. Biochem Res Commun. 1995;211:123–129. doi: 10.1006/bbrc.1995.1786. [DOI] [PubMed] [Google Scholar]
- Ting-Berreth SA, Chuong CM. Sonic Hedgehog in feather morphogenesis: induction of mesenchymal condensation and association with cell death. Dev Dyn. 1996;207:157–170. doi: 10.1002/(SICI)1097-0177(199610)207:2<157::AID-AJA4>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- Torres MA, Yang-Snyder JA, Purcell SM, DeMarais AA, McGrew LL, Moon RT. Activities of Wnt 1 class of secreted signaling factors are antagonized by the Wnt 5a class and by a dominant negative cadherin in early Xenopus development. J Cell Biol. 1996;133:1123–1137. doi: 10.1083/jcb.133.5.1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner J, Schmidt C, Nikowits W, Jr, Christ B. Compartmentalization of the somite and myogenesis in chick embryos are influenced by wnt expression. Dev Biol. 2000;228:86–94. doi: 10.1006/dbio.2000.9921. [DOI] [PubMed] [Google Scholar]
- Wang HY, Malbon CC. Wnt signaling, Ca2+, and cyclic GMP: visualizing Frizzled functions. Science. 2003;300:1529–1530. doi: 10.1126/science.1085259. [DOI] [PubMed] [Google Scholar]
- Weston CR, Davis RJ. The JNK signal transduction pathway. Curr Opin Genet Dev. 2002;12:14–21. doi: 10.1016/s0959-437x(01)00258-1. [DOI] [PubMed] [Google Scholar]
- Widelitz RB, Jiang TX, Lu J, Chuong CM. Beta-catenin in epithelial morphogenesin: conversion of part of avian foot scales into feather buds with a mutated beta-catenin. Dev Biol. 2000;219:98–114. doi: 10.1006/dbio.1999.9580. [DOI] [PubMed] [Google Scholar]
- Widelitz RB, Jiang TX, Chen CW, Scott NS, Chuong CM. Wnt7a in feather morphogenesis: involvement of anterior-posterior asymmetry and proximal-distal elongation demonstrated with an in vitro reconstitution model. Development. 1999;126:2577–2587. doi: 10.1242/dev.126.12.2577. [DOI] [PubMed] [Google Scholar]
- Wodarz A, Nusse R. Mechanisms of Wnt signaling in development. Annu Rev Cell Dev Biol. 1998;14:59–88. doi: 10.1146/annurev.cellbio.14.1.59. [DOI] [PubMed] [Google Scholar]
- Yamaguchi TP, Bradley A, McMahon AP, Jones S. A Wnt5a pathway underlies outgrowth of multiple structures in the vertebrate embryo. Development. 1999;126:1211–1223. doi: 10.1242/dev.126.6.1211. [DOI] [PubMed] [Google Scholar]
- Yu M, Wu P, Widelitz RB, Chuong CM. The morphogenesis of feathers. Nature. 2002;420:308–312. doi: 10.1038/nature01196. [DOI] [PMC free article] [PubMed] [Google Scholar]