

SUMMARY
Both reward- and punishment-related stimuli are motivationally salient and attract the attention of animals. However, it remains unclear how motivational salience is processed in the brain. Here we show that both reward- and punishment-predicting stimuli elicited robust bursting of many non-cholinergic basal forebrain (BF) neurons in behaving rats. The same BF neurons also responded with similar bursting to primary reinforcement of both valences. Reinforcement responses were modulated by expectation, with surprising reinforcement eliciting stronger BF bursting. We further demonstrate that BF burst firing predicted successful detection of near-threshold stimuli. Together, our results point to the existence of a salience-encoding system independent of stimulus valence. We propose that the encoding of motivational salience by ensemble bursting of non-cholinergic BF neurons may improve behavioral performance by affecting the activity of widespread cortical circuits, and therefore represents a novel candidate mechanism for top-down attention.
INTRODUCTION
Reward and punishment are the major driving forces of goal-directed behaviors as animals strive to maximize reward and avoid punishment. Despite having opposite hedonic valences, both reward- and punishment-related stimuli enhance arousal and attract attention (Lang and Davis, 2006). As such, both reward- and punishment-related stimuli are motivationally salient. Thus, motivational salience and hedonic valence represent two distinct but closely related attributes of reward and punishment that may be encoded by the brain. While much is known about how the opposite hedonic valences are processed by different and possibly opposing valence-specific neural systems (Daw et al., 2002; Matsumoto and Hikosaka, 2007; Paton et al., 2006; Roitman et al., 2005; Schultz et al., 1997; Seymour et al., 2005; Yacubian et al., 2006), it remains unclear whether motivational salience is determined by summing the activity of valence-specific neural systems, or alternatively, as we explore here, whether motivational salience is a valid neurobiological construct in its own right, encoded by separate neural circuits?
Support for the notion that motivational salience is processed independently of hedonic valence is provided by the findings that single neurons in several brain regions respond to both reward- and punishment-related stimuli in similar fashions (Aston-Jones and Cohen, 2005; Belova et al., 2007; Richardson and DeLong, 1990; Roesch and Olson, 2004). The mammalian basal forebrain (BF), also known as nucleus basalis, represents a particularly attractive candidate to encode motivational salience. Previous studies in both monkeys (Richardson and DeLong, 1990; Wilson and Rolls, 1990) and rats (Whalen et al., 1994) have shown that subsets of BF neurons respond to both reward- and punishment-related cues, as would be expected for the encoding of motivational salience. Furthermore, BF is required for proper execution of top-down attention toward motivationally salient stimuli (Burk and Sarter, 2001; Everitt and Robbins, 1997; Muir et al., 1994; Pang et al., 1993; Voytko et al., 1994). Finally, BOLD signals in the BF region of human subjects correlate with their motivational levels in an incentive force task (Pessiglione et al., 2007).
The BF is one of the largest neuromodulatory systems in the mammalian brain (Semba, 2000; Zaborszky and Duque, 2003). In addition to its key roles in top-down attention, the BF is also important for shaping cortical activity (Buzsaki et al., 1988; Lin et al., 2006; Riekkinen et al., 1991) and plasticity (Dykes, 1997; Kilgard and Merzenich, 1998; Weinberger, 2003). While traditionally the functional roles of BF have been mostly attributed to its cholinergic (ACh) corticopetal projection (Everitt and Robbins, 1997; Wenk, 1997), the majority of BF corticopetal projections are in fact non-ACh neurons, consisting mostly of GABAergic neurons and a smaller subset of glutamatergic neurons (Gritti et al., 1997). While the functions of non-ACh BF neurons remain poorly understood (Sarter and Bruno, 2002), the demonstration that GABAergic BF cortical afferents preferentially innervate intracortical GABAergic interneurons suggests an ideal anatomical substrate for fast modulation of cortical activity through disinhibition (Freund and Meskenaite, 1992).
In support of this view, we have recently discovered a novel mechanism by which non-ACh BF neurons may transiently enhance prefrontal cortex (PFC) activity (Lin et al., 2006). In this previous study, we showed that a homogeneous population of BF neurons, which do not change their average firing rates (2-8 Hz) across wake-sleep states and thus were classified as non-ACh neurons (Lee et al., 2005), engage in spontaneous ensemble bursting events, particularly during the waking state in rats. Such ensemble bursting events likely have a strong impact on PFC activity because they are tightly coupled with transient (∼200 msec) increases in PFC gamma oscillation (30-100 Hz) power and phase-lock with low frequency (<10 Hz) PFC field potential oscillations.
Here, we tested whether motivational salience is encoded by ensemble bursting of non-ACh BF neurons in behaving rats by simultaneously recording the activity of many BF single neurons with movable multi-electrode bundles. In particular, we investigated whether motivationally salient cues predicting reward (sucrose) or punishment (quinine) in a Go/Nogo task, as well as the reward and punishment themselves, elicited bursting responses of non-ACh BF neurons. We also studied whether the same sensory cues would fail to activate BF neurons when the cues were not motivationally salient, i.e. before associative learning and after extinction training. After confirming that ensemble bursting of non-ACh BF neurons reflects the current motivational salience of sensory stimuli, we further investigated whether the presence of BF ensemble bursting improves behavioral performance in detecting auditory stimuli presented at near-threshold levels.
RESULTS
In order to study the encoding of motivational salience and to disambiguate it from the processing of hedonic valence, we employed a Go/Nogo task because sensory cues in this task can be motivationally salient and at the same time they can be associated with either reward or punishment. Specifically, we trained rats to associate three previously neutral and clearly perceptible sensory cues with either an appetitive sucrose solution (0.3M) or an aversive quinine solution (3mM), delivered through the same licking spout. Sucrose delivery was signaled by a light cue (LS) or an auditory cue (TS), while quinine delivery was signaled by a different auditory cue (TQ) (Figure 1A). By design, the cues differed in terms of their sensory modalities (visual vs. auditory), associated motor responses (Go vs. Nogo), and the hedonic valence of predicted outcomes (reward vs. punishment). The only common feature among the three cues was their motivational salience. As expected, rats learned to lick for sucrose after TS and LS (Go response, 88.1±2.5% and 87.6±2.5%, respectively, mean±s.e.m.), while correctly avoided licking after TQ (Nogo response, 67.3±4.3%) (Figure 1B). Even when rats incorrectly licked during TQ trials, their response latency (1.63±0.08 sec) was significantly longer than the response latency in correct sucrose trials (1.10±0.09 to TS and 1.04±0.05 sec to LS, paired t-test, p<10−5). These results indicate that, after training, all cues in the Go/Nogo task were motivationally salient and rats responded to each cue according to its hedonic valence.
Figure 1. The Go/Nogo task.

(A) Schematic of the Go/Nogo task. In each trial, one of three cues, TS, TQ or LS (T, tone; L, light; subscripts S/Q indicate the associated sucrose or quinine reinforcement) was randomly chosen and presented for 2 seconds. Licking within the 5-sec response window lead to delivery of the corresponding reinforcement at the 3rd-7th licks. Licking outside the response window lead to reset of the inter-trial interval (ITI) counter. Latency, time to first lick. (B) Behavioral performance in the Go/Nogo task. The probability of Go responses (black line, left-axis) and the average latency for Go responses (red line, right-axis) for each cue (mean ± s.e.m, n=4 rats, 19 sessions). Notice that for TQ trials, the correct behavioral response is Nogo.
We first assessed whether BF neurons showed bursting responses to all three motivationally salient cues in the Go/Nogo task. A total of 210 BF neurons were recorded with movable multi-electrode bundles (Supplemental Figure S1) while rats performed the task (4 rats, 19 sessions). The most common type of BF neuronal response to cues was a short latency burst (see Supplemental Figure S2 for responses of all BF neurons and Supplemental Figure S3 for definition of bursting response). When rats correctly responded to the cues, half of BF neurons (105/210) responded to the onset of all three cues with a robust and remarkably similar short spike burst, illustrated in Figure 2A (single neuron) and Figure 2B (population). This early bursting response of BF neurons stood in clear contrast with the subsequent sustained firing modulation, which was mostly excitatory for Go responses and inhibitory for Nogo responses (Figure 2B, see also Supplemental Figure S4), indicating that the two phases of responses are dissociable. In addition, the bursting latency and amplitude of the early response to different cues were highly correlated, i.e. BF neurons with earlier and stronger responses to one cue likely showed earlier and stronger responses to other cues (Figure 2C). These strong correlations suggest that bursting responses toward all cues were qualitatively similar. Thus, while the subsequent sustained response was correlated with the rats' behavioral output (Go vs. Nogo) and the hedonic valence of the expected reinforcement (reward vs. punishment), the early bursting response was consistent only with the encoding of motivational salience that is common to all three cues.
Figure 2. Motivationally salient cues elicit bursting responses of BF neurons.
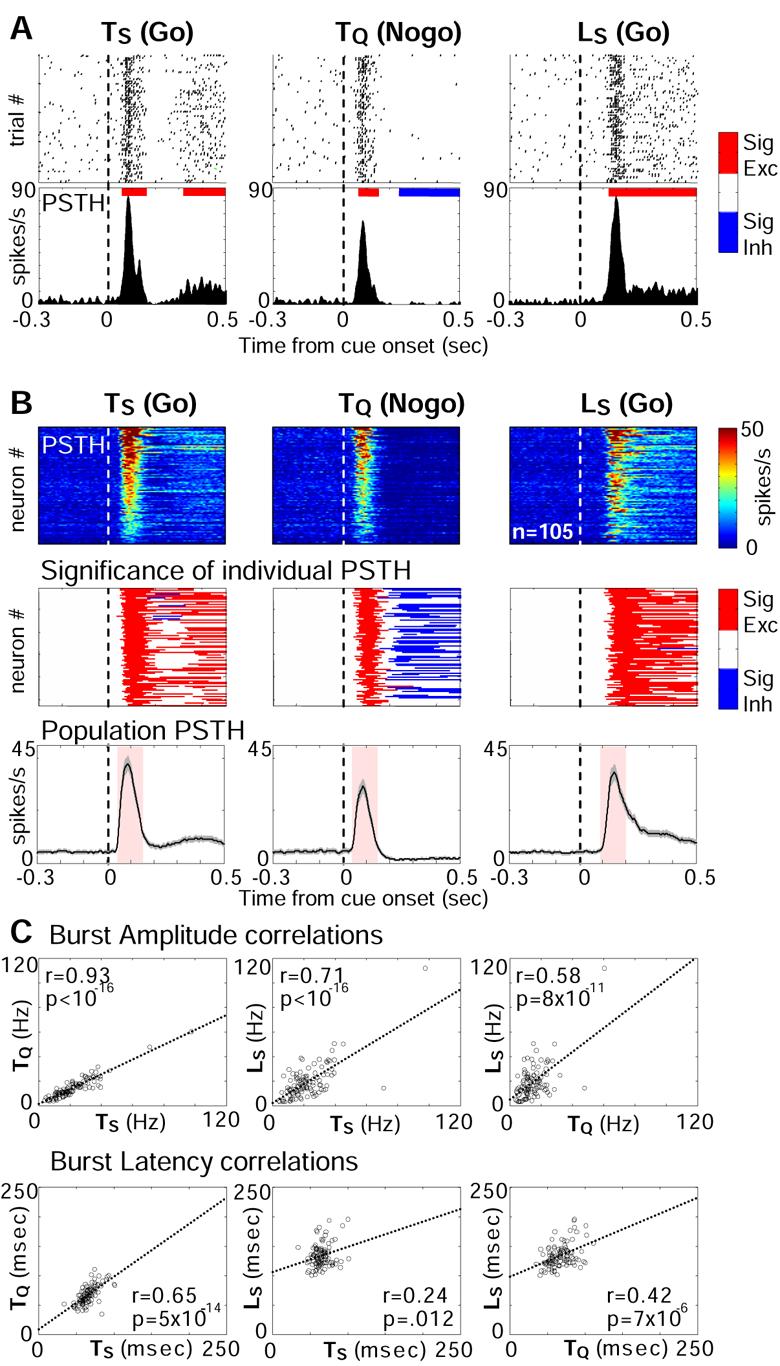
(A) Bursting responses of one BF neuron to cues when the rat made correct behavioral responses. Upper panels, raster plots aligned to cue onsets. The response latency in all trials exceeded 0.5 sec and thus was not shown. Lower panels, peri-stimulus time histogram (PSTH) of the same responses. The red and blue lines on top of the PSTHs indicated significant excitatory and inhibitory responses (p=0.005), respectively, calculated based on [−1,0] sec baseline PSTH before cue onsets. (B) BF population bursting responses to cues. Upper panels, each row represented the color-coded PSTH for one neuron. Only neurons with bursting responses to all three cues were plotted, and sorted by their burst amplitude toward TS for all subplots. Middle panels, each row represented the color-coded response significance of individual PSTHs. Lower panels, average population response to the cues (mean ± s.e.m). Pink shaded areas indicated the time windows used to calculate burst amplitude (see Experimental Procedures). (C) Correlations of burst amplitude and latency to each cue. Each circle represented one BF neuron from Figure 2B. Dotted lines were the best linear fit. The Pearson correlation coefficient and the p-values were labeled for each plot.
If BF bursting indeed encoded the abstract motivational salience of the cues, the same BF neurons should not respond to novel sensory cues that have not yet acquired motivational salience through learning. To investigate this issue, two separate groups of rats were trained on subsets of cues — one group was trained with TS and TQ but never experienced LS (novel-L group, 3 rats, 6 sessions) and the other group was trained with LS but never experienced tone cues (novel-T group, 2 rats, 4 sessions). During the recording sessions, these rats were presented with all three cues of the Go/Nogo task, as well as the same behavioral contingency. Overall, rats performed well with learned (salient) cues but rarely responded to novel (non-salient) cues (10.0±2.9% Go response) (Figures 3A and 3B). In novel-L rats, 54/100 BF neurons showed bursting responses to learned cues – TS and TQ – similar to those in Figure 2B. Yet, none of these 54 neurons showed any response to non-salient LS (Figure 3A). In novel-T rats, 58/75 BF neurons showed bursting responses to LS. Yet, only 3/58 neurons showed bursting responses to non-salient TS or TQ (Figure 3B). BF bursting responses were also absent even when the analysis was restricted to the first 10 trials rats encountered a novel cue (Supplemental Figure S5). To further demonstrate that BF bursting responses to cues was acquired through learning, one novel-L rat was subsequently trained with the LS-sucrose association. In this rat, the proportion of BF neurons with bursting responses to both TS and TQ that also responded to LS increased from 0/37 before learning to 43/44 after learning LS (Figure 3C). These results clearly show that BF bursting response to sensory cues was acquired through learning and paralleled the acquired motivational salience of the cues. The absence of responses to non-salient cues indicates that these BF neurons do not respond to sensory properties per se.
Figure 3. Absence of BF bursting responses to novel cues before learning cue-reinforcement associations.
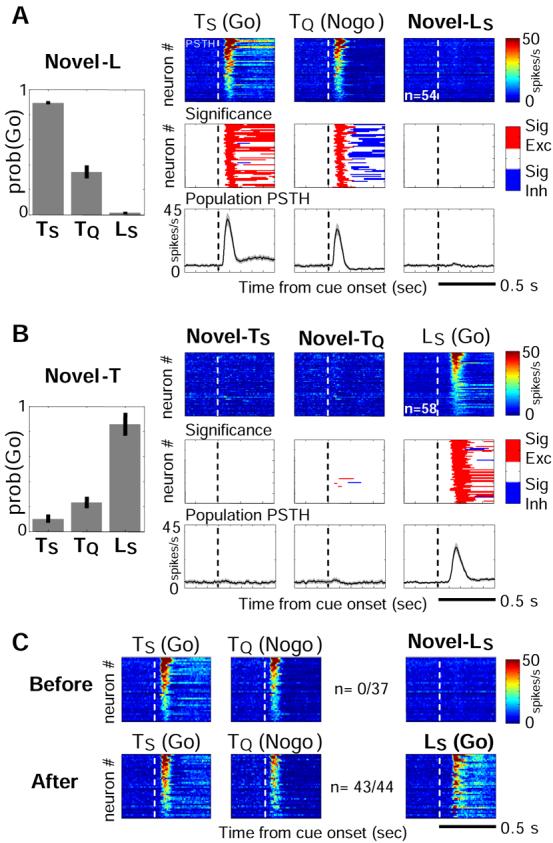
(A) Behavioral and neuronal responses to cues in novel-L rats, which had never experienced LS during training. BF neurons with bursting responses to both TS and TQ were plotted, and sorted by their burst amplitude to TS. Conventions as in Figure 2B. Notice the lack of behavioral and neuronal responses to the novel cue (LS), in contrast to prominent bursting responses to learned cues (TS and TQ). (B) Similar results for novel-T rats. (C) BF neurons recorded in one rat before (upper) and after (lower) learning LS-sucrose association. The fractions indicated the proportion of neurons with bursting response to both TS and TQ that also showed bursting response to LS.
In contrast to acquiring motivational salience through learning, sensory cues can lose their motivational salience via extinction training. We therefore investigated whether BF bursting responses to sensory cues would diminish or even disappear after extinction. Extinction training was carried out in a subset of rats that had mastered all three cue-reinforcement associations of the Go/Nogo task (3 rats, 15 sessions). In extinction sessions, rats were presented with only sucrose-predicting cues (TS and LS), but sucrose delivery was withheld following either LS (extinct-L, 6 sessions) or TS (extinct-T, 9 session). Rats maintained a high level of Go response toward rewarded cues but quickly stopped responding to cues that no longer delivered reward (23.1±4.2% Go response) (Figure 4). Despite the fact that many BF neurons continued to burst robustly toward rewarded cues (51/62 neurons in extinct-L rats, 54/67 neurons in extinct-T rats), many of these neurons did not show bursting responses toward extinguished cues (23/51 in extinct-L rats, 41/54 in extinct-T rats) (Figure 4). As a result, the population bursting amplitude to extinguished cues decreased significantly. The bursting amplitude to extinguished cues further decreased when rats consolidated their Nogo behavior (paired t-test, p<0.001) (Figure 4). Thus, even though BF bursting to extinguished cues may persist for some time after rats stopped responding to these cues, the bursting amplitude was significantly reduced. This suggests that the amplitude of BF bursting responses tracked the motivational salience of the cue.
Figure 4. Diminished BF bursting responses to cues after extinction.
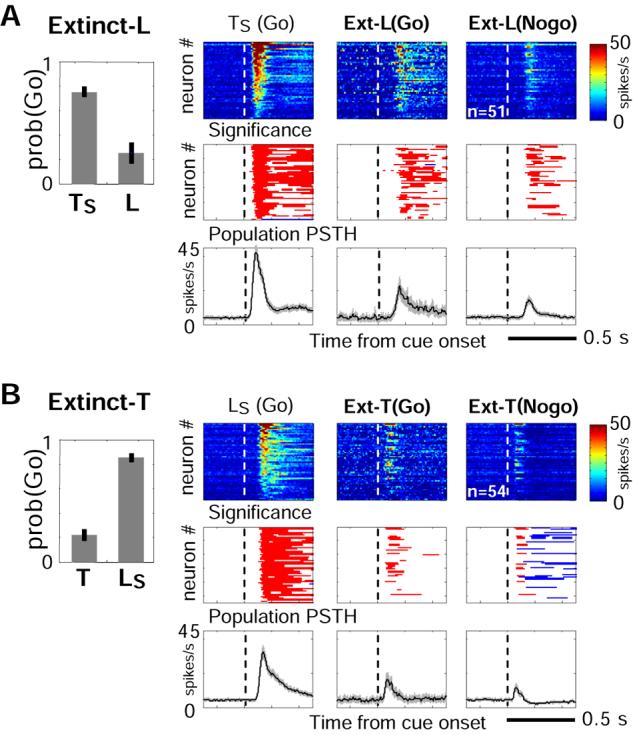
(A) Behavioral and neuronal responses to cues in extinct-L rats that underwent extinction training of the LS-sucrose association. Neurons with bursting responses to TS were plotted. Conventions as in Figure 2B. Responses to LS were plotted separately by rats' behavior responses (Go vs. Nogo). Notice the diminished neuronal responses to the extinguished cue (LS), in contrast to prominent bursting responses to the rewarded cue (TS). (B) Similar results for extinct-T rats.
Because both primary reward and punishment were motivationally salient to rats, we next investigated whether BF neurons would show similar bursting responses toward sucrose and quinine. We focused this analysis on BF neurons that encoded motivational salience of cues in the Go/Nogo task (Figure 2B). Single neuron and population responses to sucrose and quinine are shown in Figures 5A and 5B, aligned to the first delivery of sucrose and quinine in each trial. Indeed, 55% (54/99) and 69% (68/99) of BF neurons that showed bursting responses toward motivationally salient cues in the Go/Nogo task also showed significant bursting responses following the first delivery of sucrose or quinine, respectively (Figure 5B). This BF bursting response, however, was not present (or much diminished) for the first two unreinforced licks or the 2nd-5th sucrose deliveries in each trial. The bursting amplitude of individual neurons toward sucrose and quinine were highly correlated with each other, and also were correlated with burst amplitude toward sensory cues (Figure 5C). These results indicate that BF neurons also encode the motivational salience of primary reward and punishment using similar bursting responses.
Figure 5. Bursting responses of BF neurons to sucrose and quinine.

(A) Responses of the BF neuron in Figure 2A to the first delivery of sucrose or quinine in each trial. Notice that rats had to lick twice (unreinforced) before sucrose or quinine was delivered. (B) BF population bursting responses to sucrose and quinine. BF neurons with bursting responses to all three cues in Figure 2B and with at least 10 quinine trials were plotted (n=99). Significant responses were calculated based on [−1,0] sec baseline PSTH prior to cue onsets. Notice that bursting response was present only to the first, but not to subsequent 2nd-5th delivery of sucrose, nor to unreinforced licks. Pink shaded areas indicated the time windows used to calculate burst amplitude. (C) Correlations of burst amplitude to sucrose, quinine and to cues, as indicated in each plot. Red circles in left and middle panels represented neurons with significant bursting responses to sucrose or quinine. (D) BF population bursting responses to sucrose and quinine from neurons in the Novel-T group (Figure 3B). Notice that bursting responses to unexpected deliveries of sucrose and quinine following novel cues, TS and TQ (middle and right), were robust at the population level. The trial numbers were low because rats had never learned to associate TS and TQ with the delivery of tastants during training,
To further investigate whether BF bursting responses to sucrose and quinine were innate or learned, we analyzed how BF neurons in the novel-T group (Figure 3B) responded to unexpected deliveries of sucrose and quinine, on those rare occasions that rats licked within 5-sec following onsets of novel cues, TS and TQ. Despite low trial numbers, BF bursting responses to unexpected reinforcement were quite robust and their bursting amplitudes were similar (Figure 5D, paired t-test, p=0.41). This indicates that BF bursting responses to primary reward and punishment were not the result of learning but reflected the innate response property of these BF neurons. The similar bursting amplitudes toward sucrose and quinine further support the conclusion that BF encoding of motivational salience occurs independent of hedonic valence.
It is important to note that, despite the physiological heterogeneity of BF neuronal populations in vivo (Lee et al., 2004), salience-encoding BF neurons represented a homogeneous subset of cells that shared similar baseline firing rates (2-8 Hz) and peak bursting rates (30-80 Hz) (Supplemental Figure S2). These firing properties are consistent with in vitro characterizations of non-cholinergic BF neurons (Alonso et al., 1996). Moreover, unlike ACh BF neurons, which show characteristic higher firing rates during waking (WK) and rapid-eye-movement sleep (REM) than during slow-wave sleep (SWS) (Lee et al., 2005), salience-encoding BF neurons maintained similar average firing rates (2-8 Hz) between WK and SWS states (paired t-test, p>0.05) (Figure 6). On the other hand, none of the putative ACh neurons (8/143) overlapped with salience-encoding BF neurons (Supplemental Figure S6). These observations suggest that salience-encoding BF neurons are non-ACh neurons with similar firing properties to those described in our previous report (Lin et al., 2006).
Figure 6. Salience-encoding BF neurons do not change average firing rates between waking and slow-wave sleep.
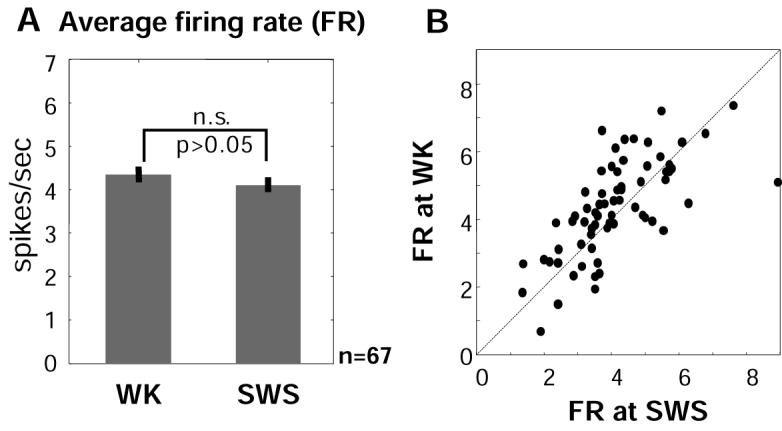
(A) Average firing rates of BF neurons (from Figure 2B) in the WK and SWS states (mean ± s.e.m.) were statistically not different. Only neurons with at least 10 minutes of SWS recording (n=67) were included in the calculation. (B) Scatter plot of the average firing rate at WK vs. SWS states. Each dot represented one BF neuron. The dashed line indicated equivalent firing rate between the two states.
To determine whether BF encoding of motivational salience has any impact on behavioral performance, we trained a separate group of three rats in a near-threshold tone-detection task. During training, rats were rewarded for making a Go response following an 80 dB tone. Importantly, during the recording phase (11 sessions), we manipulated the difficulty of tone detection by randomizing the target tone level between 56-80 dB against an additional 61 dB constant background noise. While it is possible that the quality of sound presented at different levels might differ, this manipulation created conditions in which successful tone detection could only occur on a fraction of trials when the tone sound level was near the detection threshold (Figure 7A). Thus, we were able to investigate whether successful tone detection was correlated with the presence of BF bursting response. Indeed, BF neurons displayed a clear all-or-none response pattern corresponding to successful detection — showing a clear bursting response when rats successfully detected the tone, and no response when rats failed to detect the tone (Figures 7B and 7C). Even when tones were presented near detection threshold (≤ 65 dB), the bursting response of single BF neurons was able to predict trials in which successful tone detection was achieved (Figures 7D and 7E). In other words, given the same near-threshold stimuli, successful tone detection strongly correlated with the presence of BF burst firing. Such a strong correlation suggests that the encoding of motivational salience by ensemble bursting of BF neurons may enhance behavioral performance toward attended stimuli.
Figure 7. BF bursting responses predict successful detection of near-threshold tones.

(A) Probability of detecting tones at various sound pressure levels against a 61dB background noise. Colored-dots represented data from three different rats. The dashed line indicated the logistic fit. (B) Response of a BF neuron to tone onsets, sorted by hit and miss trials (left), and then by tone sound levels (right). Bursting responses were present for hit trials but not for miss trials, regardless of tone sound levels. (C) Population responses to tone onsets in hit and miss trials for neurons with bursting responses to the 80dB tone. (D) Neuronal discrimination between hit and miss trials for near-threshold tones (≦65 dB), calculated according to signal detection theory. Each row represented the color-coded choice probability of a BF neuron from Figure 7C as a function of time (see Experimental Procedures). Only bins reaching statistical significance were plotted. (E) Distribution of the maximal choice probability in the [0.05 0.25] sec interval. 64/67 neurons in Figure 7E reached significance level.
Within successful detection trials, we also found that the target sound level modulated the latency and amplitude of BF bursting responses (repeated measure ANOVA, p<0.001) (Figures 8A and 8B). This graded bursting response likely reflected the varying levels of motivational salience for different tone levels, with louder tones associated with higher motivational salience. Consistent with this interpretation, we found that in trials with stronger BF bursting response to the target, and hence higher motivational salience, the behavioral response latency was shorter (Figure 8D). Intriguingly, BF response amplitudes to the target tone and the water reward were inversely correlated: stronger BF bursting responses to the target lead to weaker BF responses to the reward, and vise versa (Figure 8A). This pattern was also evident at the single neuron level (Figures 8B and 8C). Overall, this anti-correlated response pattern supports the notion that expected rewards are less salient, while surprising ones are more salient.
Figure 8. Graded BF bursting responses in the detection task.

(A) Average population response to the tones and reward delivery in the detection task, sorted by tone sound levels. Notice that stronger bursting responses to target tones were associated with weaker responses to reward delivery, and vise versa. (B) Normalized burst amplitude to tones (red) and reward delivery (blue) (mean ± s.e.m, n=67). Burst amplitude of each BF neuron at each condition was normalized to its burst amplitude toward 80 dB tone (hit trials). (C) Population responses to the water reward for the same BF neurons as in Figure 7C. Notice that the bursting response is temporally broader than those in Figure 5, which likely reflects the slight temporal jitter between nose poke and licking for water reward in this experiment. (D) Mean behavioral response latency in hit trials, sorted by tone sound levels (mean ± s.e.m, n=3 rats, 11 sessions). Notice that stronger bursting responses to target tones were associated with faster response latencies.
DISCUSSION
This study examined whether BF neurons encode motivational salience of attended stimuli in behaving rats. Our main conclusion is that motivational salience is encoded by phasic bursting of non-ACh BF neurons. We found that many BF neurons showed robust bursting responses to motivationally salient sensory cues that reliably predicted reinforcement, irrespective of their sensory modalities (visual or auditory), associated motor responses (Go or Nogo) or hedonic valences (reward or punishment) (Figure 2). However, BF bursting was absent (or much diminished) when the same sensory cues were not motivationally salient, i.e. before associative learning (Figure 3) or after extinction (Figure 4). The same BF neurons also responded with similar bursting to innately salient primary reinforcement (sucrose and quinine, Figure 5). The response amplitude to reinforcement was modulated by expectation, with surprising reinforcement eliciting stronger BF bursting (Figure 8). We further demonstrated that the encoding of motivational salience by BF bursting predicted successful detection of near-threshold tones (Figure 7). Finally, these salience-encoding BF neurons were likely non-ACh neurons because, differently from typical ACh BF neurons, they did not change their average firing rates across wake-sleep states (Lee et al., 2005) (Figure 6). Together, these results establish that ensemble bursting of non-ACh BF neurons encodes motivational salience, which may improve behavioral performance toward motivationally salient stimuli.
What is the role of BF bursting in detecting near-threshold stimuli?
When rats successfully detected the target tone in the near-threshold detection task, the target sound level modulated the latency and amplitude of BF bursting responses (Figures 8A and 8B). One interpretation of this apparent correlation is that BF ensemble bursting may reflect the sensory detection process. However, as we demonstrated in Figure 3, sensory cues that were clearly perceptible did not elicit any BF response before associative learning. The lack of BF response in this scenario indicates that sensory detection can occur without recruiting BF ensemble bursting. Therefore, BF bursting very likely does not reflect sensory detection.
It is important to recognize that, as rats learned to associate the target tone with reward, active reporting of tone detection (with nose poke and licking) required not only sensory detection but also additional cognitive processes. While BF bursting likely does not reflect sensory detection, it may play an essential role in subsequent cognitive processes via encoding motivational salience. Therefore, the graded BF bursting response in the detection task likely represents different levels of motivational salience associated with distinct tones. Louder tones that were successfully detected were associated with higher motivational salience, perhaps reflecting the higher confidence level of tone detection. Higher motivational salience, in turn, was associated with faster response latency (Figure 8D).
The tight correlation between BF bursting and successful tone detection (Figure 7) suggests that BF encoding of motivational salience may play a causal role to enhance behavioral performance toward attended stimuli. In support of this causal view, inactivation or excitotoxic lesions of the BF lead to impairments in detecting briefly presented stimuli (Burk and Sarter, 2001; Muir et al., 1994; Pang et al., 1993). The causal view is also consistent with BF bursting occurring early in the information processing stream (50 msec for auditory and 100 msec for visual stimuli), allowing the BF to provide powerful influences on cortical activity (Lin et al., 2006). Therefore, BF bursting may serve to enhance the cortical representation of the detected stimuli for the purpose of reinforcement-guided behaviors.
Expectation modulates BF bursting responses to reinforcement
The motivational salience of reinforcement depends on expectation: well-predicted reinforcement is less salient while surprising ones are more salient. Modulation of salience levels by expectation likely accounted for the varying BF bursting amplitudes observed toward reinforcement. In the Go/Nogo task, BF responses to surprising reinforcement following novel cues (Figure 5D, middle and right) were greater than responses to expected reinforcement following the well learned cue (Figure 5D, left, paired t-test, p<0.001). The larger response to quinine than to sucrose in Figure 5B (paired t-test, p<0.001) likely reflects the fact that quinine occurred when rats made incorrect Go responses, and thus more surprising, while sucrose was well predicted by the cue.
The same logic also applies to BF responses to the water reward in the tone detection task (Figure 8). When the target stimulus was more salient, the reward was better predicted. Therefore once reward was actually received, it was less salient and elicited a smaller bursting response. On the other hand, target stimuli presented near threshold level were less salient and resulted in poorer predictions of reward. In this case, reward delivery was more surprising and salient, and elicited a stronger bursting response.
Similar effects of expectation have been described in the amygdala (Belova et al., 2007). These authors showed that amygdala neurons respond to reward and/or aversive air-puff. In subsets of amygdala neurons, the reinforcement responses are similarly modulated by expectation. Since amygdala is one of the major input structures of the BF (Jolkkonen et al., 2002), these similar response properties in the amygdala are likely relayed to the BF for the computation of motivational salience.
The functional significance of BF sustained responses following initial bursting
In addition to the initial bursting that encodes motivational salience, BF neurons also displayed subsequent sustained response that correlates with stimulus valence and/or motor responses, i.e. excitatory in correct Go trials and inhibitory in correct Nogo trials (Figure 2B). This correlation extended to error Go trials following TQ presentations. In these trials, BF neurons responded with similar initial bursting, followed by neither excitation nor inhibition during the sustained response phase, at intermediate amplitudes between those of correct Go trials and correct Nogo trials (Supplemental Figure S4). In parallel, motor responses in error Go trials were also intermediate between the other two trial types, showing significantly slower response latency relative to correct Go trials (Figure 1B). Therefore, the intermediate sustained response in error Go trials may reflect the ambivalent valence estimation and/or the sluggish motor response. It is unclear at this point whether this correlation reflects a causal relationship.
Our results support the notion that the initial bursting and subsequent sustained BF responses represent two dissociable neural processes encoding different information. It is likely that the valence (or motor) information conveyed by sustained response is not computed locally within the BF, because similar initial bursting can be followed by different and opposite patterns of sustained response. Instead, this valence information is likely relayed from other valence-specific neural systems to the BF, such as midbrain dopaminergic neurons.
Separate encoding of motivational salience and hedonic valence
Our results provide clear evidence that motivational salience and hedonic valence, two distinct but closely related attributes of reward and punishment, are separately encoded in the brain. In this context, our findings add to the growing list of brain regions that may encode motivational salience, including the locus coeruleus (LC), amygdala, premotor cortex and the BF (Aston-Jones and Cohen, 2005; Belova et al., 2007; Richardson and DeLong, 1990; Roesch and Olson, 2004), and further highlight important differences on how motivational salience is encoded in different brain structures. In particular, BF encoding of motivational salience utilizes a highly homogeneous bursting response that is not seen in the amygdala and the premotor cortex. In addition, the fast bursting activity of BF neurons may broadcast the salience information via their widespread corticopetal projections to influence salience-related activity in cortical networks (Lin et al., 2006).
One potential concern of our interpretation is that BF neurons, in addition to encoding motivational salience, may also encode valence information. In particular, the consistently larger BF bursting amplitude in TS trials than in TQ trials of the Go/Nogo task (Figure 2B, paired t-test, p<0.001) raises the possibility that valence information may be encoded by the differential response to the two stimuli. This interpretation, however, is inconsistent with the finding that variations in BF bursting amplitude can be used to signal different levels of motivational salience, as we demonstrated in the detection task (Figure 8). Therefore, the different bursting amplitudes toward TS and TQ most likely reflected different levels of salience rather than signaling opposite valences.
BF encoding of motivational salient reported here bears a close resemblance to how noradrenergic neurons in the LC respond to salient target stimuli with phasic bursting responses at short latency, and with little or no response to non-salient stimuli (Aston-Jones and Cohen, 2005). However, the two systems likely differ in the timing of their respective bursting responses. While LC phasic activation is better time-locked to motor responses, BF ensemble bursting is time-locked to stimulus onset. The stimulus-locking property of BF bursting is supported by the similar burst latency to TS and TQ (Figure 2C, paired t-test, p>0.05), despite the qualitatively different motor responses in these trial types. Therefore, in the presence of motivationally salient stimuli, ensemble bursting of BF non-ACh neurons may occur earlier than the phasic response of LC neurons.
Another interesting comparison with the non-ACh BF neurons is midbrain dopaminergic neurons, which encode reward prediction error signals with phasic bursting responses (Schultz et al., 1997). Our results suggest that the brain utilizes these two major neuromodulatory systems, with similar bursting responses at similar latencies, to separately encode the motivational salience and hedonic valence of attended stimuli. One consequence of this parallel is that both systems will be simultaneously activated by reward-related stimuli, both capable of powerfully modulating cortical activity. It is therefore important to delineate the respective contributions of these subcortical salience and valence systems in shaping reward-related cortical activity.
BF ensemble bursting as a candidate mechanism for top-down attention
Animals are faced with the constant challenge of deciding which sensory stimuli should receive their attention. In this regard, motivational salience is closely tied with attention, since both reward- and punishment-related stimuli represent a special subset of stimuli that attract the animals' attention. Hence, we referred to this aspect of attention as “top-down attention”, to reflect the intuition that evaluating the motivational salience associated with a sensory stimulus requires not only sensory detection but also cognitive evaluation based on past experience.
Ensemble bursting of non-ACh BF neurons shares several common features with top-down attention. First, encoding the motivational salience of a stimulus represents a critical step in determining whether and when animals should attend to incoming sensory information. Second, similar to how top-down attention improves behavioral performance, BF bursting was found to be tightly correlated with successful detection in a tone detection task. As we discussed above, BF bursting may play a causal, not just correlational, role in improving behavioral performance. Third, the influences of top-down attention on cortical processing to transiently enhance gamma oscillations and event-related potentials (ERPs) (Engel et al., 2001; Herrmann and Knight, 2001; Ward, 2003) are closely paralleled by the ability of BF ensemble bursting to transiently enhance cortical gamma oscillation power and to phase-lock with cortical local field potentials (LFPs) (Lin et al., 2006).
These similarities therefore suggest the novel hypothesis that ensemble bursting of non-ACh BF neurons, via their influences on cortical networks, may transform the encoding of motivational salience into transient amplification of cortical activity, thereby mediating the influences of top-down attention on neural activity and behavior. Lending further support to this hypothesis, BF ensemble bursting likely exerts its maximal impact on cortical activity at 50-300 msec following stimulus onsets, estimated based on BF bursting latency (50-100 msec) and the duration of BF-mediated fast cortical modulation (∼200 msec). This time window matches the time window of induced gamma oscillation and the N2/P3 components of ERPs, both of which represent reliable indicators of attentional modulation on stimulus detection, demonstrated in a wide range of behavioral tasks (Engel et al., 2001; Folstein and Van Petten, 2008; Herrmann and Knight, 2001; Sambeth et al., 2003; Ward, 2003).
It is important to note that attention is not a monolithic mechanism but rather a collection of processes. Therefore, it is likely that BF ensemble bursting may underlie only certain aspects of attention. In particular, we find that the conceptualization of ‘attention for performance’ (Holland and Gallagher, 1999) best captures the type of attention BF ensemble bursting may embody. Holland and Gallagher distinguished between two aspects of attention in associative learning: attention for performance vs. attention for learning, both aspects are supported by amygdala-BF interactions (Holland, 2007; Maddux et al., 2007). Attention for learning affects the acquisition of new learning, while attention for performance is involved in modulating current actions on well-learned tasks. Under the framework of attention for performance, consistent predictors of motivationally salient outcomes command more attention and are more likely to control action than less consistent predictors. Therefore, the level of attention would be higher for tones eliciting stronger BF bursting responses in the detection task, because stimuli with higher motivational salience are better predictors of reinforcement than less salient ones. Together, these observations support the hypothesis that top-down attention, in particular the concept of attention for performance, may be mediated by fast subcortical modulation, originating from non-ACh BF neurons.
The contributions of non-ACh and ACh BF neurons to attention
The roles of BF in attention are well supported by BF lesion experiments, which lead to selective impairments in attention (Burk and Sarter, 2001; Muir et al., 1994; Voytko et al., 1994). While traditionally the roles in attention have been mostly attributed to the ACh corticopetal projection (Everitt and Robbins, 1997; Wenk, 1997), selective immunotoxic lesions of ACh BF neurons generally lead to less severe behavioral impairments than excitotoxic lesions of the BF (Burk and Sarter, 2001; McGaughy et al., 2002; Muir et al., 1994). These observations indirectly support the notion that non-ACh BF neurons, which are preferentially targeted by excitotoxic lesions (Page et al. 1991), may also play important roles in attention (Sarter and Bruno, 2002). The current study, therefore, provides the first direct neurophysiological evidence that the anatomically prominent, but functionally poorly understood, non-ACh BF corticopetal systems play an important role in encoding motivational salience and in mediating some aspects of top-down attention.
Here, the non-ACh identity of salience-encoding BF neurons was established by comparing their firing characteristics to in vitro characterizations of non-ACh BF corticopetal neurons (Alonso et al., 1996), and by demonstrating that salience-encoding BF neurons did not change average firing rates across wake-sleep states, as would be expected from ACh neurons (Lee et al., 2005). BF neurons with state-dependent firing modulations compatible with those of ACh neurons were also observed in our data set in a small subset of neurons, which did not overlap with salience-encoding BF neurons (Supplemental Figure S6). However, given the small sample size and without independent means of verifying their neurochemical identity, we are unable to determine how ACh BF neurons behave in the Go/Nogo task.
To reconcile with the vast literature demonstrating an essential role of ACh BF neurons in attention (Everitt and Robbins, 1997; Wenk, 1997), we propose that ACh and non-ACh BF neurons may represent two parallel attention systems, perhaps serving complementary roles by operating on different temporal scales. Non-ACh BF neurons, as discussed earlier, are likely responsible for fast effects of attention within the first 300 msec of stimulus onsets, and closely associated with attention for performance. On the other hand, ACh modulations are much slower, constrained by the slower-acting metabotropic muscarinic receptors (Kaczmarek and Levitan, 1987) as well as the slow temporal dynamics of cortical ACh concentration, ranging from ACh transients lasting 2-4 seconds (Parikh et al., 2007) to the time scale of arousal states (minutes to hours). Based on the importance of ACh modulation in cortical plasticity (Dykes, 1997; Kilgard and Merzenich, 1998; Weinberger, 2003), ACh modulation may be more important for learning. Therefore, ACh modulation likely mediates slower forms of attention on time scales of seconds to minutes, such as sustained attention and arousal, and better linked with attention for learning (Maddux et al., 2007).
EXPERIMENTAL PROCEDURES
Surgery and electrode design
All procedures were approved by the Duke IACUC and performed in accordance with NIH guidelines. Adult male Long-Evans rats, 350-450 gm, were used in this study. Detailed surgical procedures have been previously described (Lin et al., 2006; Nicolelis et al., 1997). To record BF single unit activities in behaving rats while minimizing damage to overlying brain regions, we developed a movable array that incorporated 32 Isonel-coated tungsten microwire electrodes (35μm diameter, impedance 0.2-0.5 MΩmeasured at 1kHz, 50nA) into 4 movable bundles. Each bundle contained eight microwires and was ensheathed in a 29-gauge stainless steel cannula and controlled by precision microdrives (Supplemental Figure S1A). The four cannulae were precisely positioned to target the BF on both hemispheres at AP −0.25mm, ML ±2.0mm and AP −1.0mm, ML ±2.5mm relative to Bregma (Paxinos and Watson, 2005). During surgery, the cannulae were lowered to DV 6.5mm below the pial surface, and the electrodes were advanced 0.75mm from the tip of cannulae. Rats were allowed 10-14 days to recover from surgery. Electrodes were advanced 125μm after each recording session to sample BF neurons between DV 7.25-8.25 mm. Cannulae and electrode tip locations were verified with cresyl violet staining of histological sections after the end of the experiment. All electrode arrays were found at expected positions.
Behavioral training
Behavioral apparatus
The behavioral chambers (30.5 cm × 24.1cm × 21.0 cm) were constructed with plexiglass walls to minimize electrical noise and placed inside sound and light attenuated boxes. Infrared (IR) illumination was used while rats' behaviors were videotaped. A custom-built multi-barrel sipper tube was used to deliver sucrose (0.3M) or quinine hydrochloride (3mM) solutions, controlled by separate solenoid valves. The fluid delivery system was pressurized at 2.6 psi such that each solenoid opening (10 msec) delivered 10μL of solution. An IR photo-beam lickometer, capable of registering the timing of single licks with 5 msec temporal resolution, was positioned in front of the sipper tube. Each delivery of solution was always triggered by the tongue breaking the IR beam, thus ensuring fluid delivery directly onto the tongue.
Stimulus delivery and calibration
Each chamber was equipped with a house light (1.12 W) and 2 speakers (ENV-224AM, 224BM, Med Associates Inc, VT). The ENV-224 AM speaker has a flat output response of 1.0 to 7.0 KHz (at 700 Hz and 7.5 KHz the audio signal is approximately 10 dB down), which was used to deliver pure tone target stimuli. The ENV-224BM speaker has a flat output from 5 to 15 KHz (at 2 KHz and 18 KHz the audio signal is approximately 10 dB down), which was used to present the constant 61 dB white noise in the detection task. The rise and fall times of all stimuli were set at 3 msec. Both speaker outputs were calibrated before and after the experiment with a digital sound level meter (Radio Shack, CAT. No. 33-2055) while emitting 60 dB and 80 dB white noise (SPL, A-weighting). The difference of sound levels at different corners of the box was less than 2dB.
Water deprivation
Rats were water deprived and weighed daily to maintain their body weight at 90% of their original weight. They were allowed access to water for 30 minutes per day in their home cage after the daily training session had ended for 1-2 hours. In addition, they were given free water access one day per week. Water restriction was discontinued for at least 3 days in preparation for surgery and 2 weeks for post-surgery recovery.
Behavioral training of the Go/Nogo task
A schematic of the task is depicted in Figure 1A. The beginning of each behavioral session was signaled by turning on the house light. Initially, rats were trained to lick with a variable interval (20 sec) reinforcement schedule to obtain five drops of sucrose reward. In the following sessions, rats learned three cue-reinforcement associations in successive phases: TS-sucrose, TQ -quinine and LS-sucrose. TS and TQ were tones at either 6 or 10 kHz (80 dB, 2 sec duration, counter-balanced across animals), and LS was the house light turned off for 2 sec. One cue-reinforcement association was added in each phase in the order of TS, TQ and LS. On each trial, a cue was randomly selected and the inter-trial interval (ITI) was pseudo-randomly chosen between 6-18 seconds. The onset of the cue signaled a 5-sec window during which licking led to delivery of sucrose or quinine, immediately following the 3rd to the 7th lick. The first two licks in each trial were not reinforced to dissociate the motor aspect of licking from consumption of sucrose or quinine. To reduce false-alarm licking responses in the absence of cues, the ITI counter was reset every time licking was detected outside the 5-sec response window. Rats were allowed to roam freely within the behavioral chamber and were not required to respond in each trial. Each daily training session lasted 60-90min. Each training phase was complete when rats performed more than 2.5 correct trials (hits) per minute for 3 consecutive days, indicating a high hit rate and a low false-alarm rate. To prevent excessive aversion to quinine consumption early in the training process, 1mM quinine was used for initial training sessions and subsequently replaced by 3mM quinine as behavioral performance improved. TS and TQ were deliberately chosen to be similar in order to increase task difficulty such that there were enough error trials following TQ for the analysis of BF neuronal responses toward quinine.
Behavioral training for Novel-T / Novel-L groups
The training procedures were the same as in the Go/Nogo task, except that rats were only trained with subset of cues during the training phase. Rats in the Novel-T group only learned LS but never experienced TS or TQ. Rats in the Novel-L group only learned TS and TQ but never experienced LS. At the completion of training, rats underwent surgery for electrode implantation. During the recording phase, all three cues were presented to the rats with the same contingencies as in the Go/Nogo task. To ensure that rats did not learn the novel cues during the recording phase, each rat was recorded for only 1-3 sessions under this condition.
Behavioral training for Extinct-T / Extinct-L groups
Rats that have been trained and recorded in the Go/Nogo task were subsequently recorded under extinction training for one of the two sucrose-predicting cues.
Near-threshold tone detection task
A separate group of rats were trained with TS (6 kHz, 80 dB, 2 sec)-water association using procedures described in the Go/Nogo task with the following modifications. An IR nose poke unit was used instead of the photobeam lickometer. Once nose poking was detected within the 5-sec response window, water (50μL) was delivered from the end of the nose poke as the reward. After rats acquired this association, they underwent surgery for BF electrode implantation. During recording sessions, the target sound level in each trial was randomly drawn from a list of possible levels between 56-80 dB (without replacement). An additional constant 61dB noise was delivered by the second speaker to increase detection difficulty. In order to encourage the use of previously learned behavioral contingency and to maintain a high level of performance, the trials were divided into 80 dB (30%), 70 dB (20%), 65 dB (20%), 62 dB (20%) and 60 dB (10%).The target sound levels were sometimes adjusted according to the behavioral performance of the rat. Randomization of the target sound level as well as the ITI minimized the effect of expectation.
Recording and spike sorting
Recording started with the behavioral session that lasted 60-90 minutes. After the completion of the behavioral session, rats were sometimes recorded for 2-3 additional hours in the recording chamber without reward or any behavioral contingency. Most rats would rest and undergo their natural wake-sleep cycle during this period, which allowed us to investigate the firing rate modulations of the same BF neurons in various wake-sleep states. At the conclusion of each recording session, BF electrodes were advanced 125μm and 3-7 days elapsed before the next recording session.
Electrical signals collected from each electrode were referenced to a common skull screw, differentially filtered for single unit activity (154 - 8.8k Hz) and LFPs (0.4 - 240 Hz), and recorded using a Multichannel Acquisition Processor (MAP, Plexon Inc, Dallas, TX) as previously described (Nicolelis et al., 2003). Single unit isolation is of paramount importance to our study in order to clearly separate ACh from non-ACh neurons. During online sorting, only units with at least 3:1 signal-to-noise ratio were selected. Waveforms crossing a threshold level were recorded for offline spike sorting using OfflineSorter (Plexon Inc, Dallas, TX). Spike sorting was based on various features of the waveform, including the first three principal components (PCs) of spike waveforms, peak and valley amplitude of each waveform, and waveform amplitude at particular time points. Inter-spike interval histogram was further used to ensure that a minimal number of spikes (<0.1%) occurred within the action potential refractory period (set at 1.5 msec). Only single units with clear separation from the noise cluster were used for further analyses. Detailed spike sorting method, as well as quantification of cluster separation, have been previously described (Nicolelis et al., 2003).
Data analyses
Identification of wake-sleep states
The three major wake-sleep states: WK, SWS and REM, were determined using the state space method we previously developed (Gervasoni et al., 2004). Each dot in the 2-D state-space was calculated based on 1-sec of LFP data, represented by two spectral amplitude ratios 0.5-4.5 / 0.5-9 Hz (x-axis) and 0.5-20 / 0.5-55 Hz (y-axis). Points within the three main clusters were labeled as the three major states, while points in between clusters were labeled as state transitions (Supplemental Figure S1D). Identification of wake-sleep states allowed us to investigate the firing rate modulations of the same BF neurons in various wake-sleep states. Furthermore, to ensure that behavioral performances and neuronal responses were calculated based only on cues presented during WK, sensory stimuli not presented during the WK state were removed from further analysis.
Significance of PSTH modulations
Significant responses of each BF neuron to all behavioral events were determined using the method developed by Wiest et al (Wiest et al., 2005). Briefly, the statistical significance of PSTHs (binned at 1 msec) was determined by comparing cumulative frequency histograms (CFHs) of PSTHs after cue onsets against the cumulative sum distribution of baseline PSTH before cue onsets ([−1, 0]s), estimated based on 1000 bootstrapped samples (with replacement). The response onset latency was defined as the first bin in which post-cue CFH exceeded the cumulative sum distribution from the baseline PSTH (p=0.01, two-sided). The response offset was identified as the first zero crossing of the derivative of the cumulative deviation from the baseline cumulative sum distribution, meaning that the post-cue CFH was no longer deviating from its expected growth with time. The magnitude of the response was quantified as the number of excess spikes between onset and offset, compared with the baseline expected number. After the offset of each significant response, the post-cue CFH was reset to zero in order to detect subsequent significant responses in the post- cue PSTH. A minimum response amplitude of 0.3 spike (per response) was required to be considered a significant response. To determine the significance of sucrose and quinine responses, we chose to compare against baseline PSTHs before cue onsets ([−1, 0]s) where the firing rate was stable.
Definition of bursting responses
The distributions of onset latency for excitatory and inhibitory responses for various behavioral events are shown in Supplemental Figure S3. Most BF neurons responded to salient events with a robust short latency excitatory response, which we defined as bursting responses. Specifically, bursting responses were excitatory responses with latencies between 20-120 msec (for TS, TQ, sucrose and quinine) or between 80-200 msec (for LS) (gray shaded areas in Supplemental Figures S3A and S3B). We also defined the amplitude of bursting responses as the mean firing rate in the 40-150 msec window (for TS, TQ, sucrose and quinine) or the 90-200 msec window (for LS), with the average baseline firing rate ([−1, 0]s before cue onsets) subtracted. These windows were indicated by the pink shaded areas in Figures 2B and 5B. The time window for calculating bursting amplitude toward the water reward in the detection task was expanded to 50-250 msec to account for the temporal jitter between nose poke and the actual licking.
State-dependent firing rates
Only salience-encoding BF neurons (from Figure 2B) with at least 10 minutes of SWS recording in the Go/Nogo task (n=67) were included in the calculation. The firing rate during WK was calculated by averaging firing rate at [−1, 0] sec before cue onsets. The firing rate during SWS was calculated by dividing total spike counts during SWS by the duration of SWS. Psychometric functions for tone detection were constructed by plotting the percent of Go responses against tone intensity levels. Data points were regressed with a logistic function.
Neuronal discrimination of successful tone detection
Signal detection theory (Green and Swets, 1966) was used to quantify neuronal discrimination of successful (hit) versus failed (miss) tone detections. Receiver Operating Characteristic (ROC) curves, in particular the area under curve (AUC) measure, was used to determine how neuronal responses in hit trials and miss trials differed. The AUC measure, referred to as choice probability in Figures 7D and 7E, quantified the difference of the number of spikes within a 100 msec window for one BF neuron on hit trials vs. miss trials when tones were presented at near-threshold levels (≤65dB). A choice probability of 0.5 represents a complete overlap between the two distributions, while a choice probability of 1 (or 0) indicates complete non-overlap between the two distributions, with one distribution having larger (or smaller) values. The statistical significance of the choice probability was established by comparing against a null distribution of choice probability values generated by randomly permuting the identity of hit and miss trials 1000 times (p<0.01, two-sided). The choice probability (and its statistical significance) was generated for each BF neuron, at time lags between [−1, 1] sec of tone onsets with 10 msec steps. Therefore, a significant choice probability (>0.5) for a BF neuron at a specific time lag indicated that the neuronal response at that time lag (within a 100 msec window) was stronger for hit trials compared to miss trials.
Supplementary Material
ACKNOWLEDGEMENTS
We thank A. Oliveira-Maia, I. Davison, M. C. Wiest and R. Costa for critical discussions and comments on the manuscript, J. Meloy for technical assistance. S.L. is a recipient of NARSAD 2008 Young Investigator Award. This research was supported by NIH grants R01DE011451 (NIDCR) and P50MH060451 (NIMH) to M.A.L.N. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIDCR, NIMH or NIH.
Footnotes
The Supplemental Data for this article can be found online at http://www.neuron.org/cgi/
REFERENCES
- Alonso A, Khateb A, Fort P, Jones BE, Muhlethaler M. Differential oscillatory properties of cholinergic and noncholinergic nucleus basalis neurons in guinea pig brain slice. Eur J Neurosci. 1996;8:169–182. doi: 10.1111/j.1460-9568.1996.tb01178.x. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–450. doi: 10.1146/annurev.neuro.28.061604.135709. [DOI] [PubMed] [Google Scholar]
- Belova MA, Paton JJ, Morrison SE, Salzman CD. Expectation modulates neural responses to pleasant and aversive stimuli in primate amygdala. Neuron. 2007;55:970–984. doi: 10.1016/j.neuron.2007.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burk JA, Sarter M. Dissociation between the attentional functions mediated via basal forebrain cholinergic and GABAergic neurons. Neuroscience. 2001;105:899–909. doi: 10.1016/s0306-4522(01)00233-0. [DOI] [PubMed] [Google Scholar]
- Buzsaki G, Bickford R, Ponomareff G, Thal L, Mandel R, Gage F. Nucleus basalis and thalamic control of neocortical activity in the freely moving rat. J. Neurosci. 1988;8:4007–4026. doi: 10.1523/JNEUROSCI.08-11-04007.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daw ND, Kakade S, Dayan P. Opponent interactions between serotonin and dopamine. Neural Netw. 2002;15:603–616. doi: 10.1016/s0893-6080(02)00052-7. [DOI] [PubMed] [Google Scholar]
- Dykes RW. Mechanisms controlling neuronal plasticity in somatosensory cortex. Can J Physiol Pharmacol. 1997;75:535–545. [PubMed] [Google Scholar]
- Engel AK, Fries P, Singer W. Dynamic predictions: oscillations and synchrony in top-down processing. Nat Rev Neurosci. 2001;2:704–716. doi: 10.1038/35094565. [DOI] [PubMed] [Google Scholar]
- Everitt BJ, Robbins TW. Central cholinergic systems and cognition. Annu Rev Psychol. 1997;48:649–684. doi: 10.1146/annurev.psych.48.1.649. [DOI] [PubMed] [Google Scholar]
- Folstein JR, Van Petten C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 2008;45:152–170. doi: 10.1111/j.1469-8986.2007.00602.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freund TF, Meskenaite V. gamma-Aminobutyric acid-containing basal forebrain neurons innervate inhibitory interneurons in the neocortex. Proc Natl Acad Sci U S A. 1992;89:738–742. doi: 10.1073/pnas.89.2.738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gervasoni D, Lin SC, Ribeiro S, Soares ES, Pantoja J, Nicolelis MA. Global forebrain dynamics predict rat behavioral states and their transitions. J Neurosci. 2004;24:11137–11147. doi: 10.1523/JNEUROSCI.3524-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green DM, Swets JA. Signal detection theory and psychophysics. John Wiley; New York: 1966. [Google Scholar]
- Gritti I, Mainville L, Mancia M, Jones BE. GABAergic and other noncholinergic basal forebrain neurons, together with cholinergic neurons, project to the mesocortex and isocortex in the rat. J Comp Neurol. 1997;383:163–177. [PubMed] [Google Scholar]
- Herrmann CS, Knight RT. Mechanisms of human attention: event-related potentials and oscillations. Neurosci Biobehav Rev. 2001;25:465–476. doi: 10.1016/s0149-7634(01)00027-6. [DOI] [PubMed] [Google Scholar]
- Holland PC. Disconnection of the amygdala central nucleus and the substantia innominata/nucleus basalis magnocellularis disrupts performance in a sustained attention task. Behav Neurosci. 2007;121:80–89. doi: 10.1037/0735-7044.121.1.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holland PC, Gallagher M. Amygdala circuitry in attentional and representational processes. Trends in Cognitive Sciences. 1999;3:65–73. doi: 10.1016/s1364-6613(98)01271-6. [DOI] [PubMed] [Google Scholar]
- Jolkkonen E, Miettinen R, Pikkarainen M, Pitkanen A. Projections from the amygdaloid complex to the magnocellular cholinergic basal forebrain in rat. Neuroscience. 2002;111:133–149. doi: 10.1016/s0306-4522(01)00578-4. [DOI] [PubMed] [Google Scholar]
- Kaczmarek LK, Levitan IB. Neuromodulation: The Biochemical Control of Neuronal Excitability. Oxford Univ. Press; New York: 1987. [Google Scholar]
- Kilgard MP, Merzenich MM. Cortical map reorganization enabled by nucleus basalis activity. Science. 1998;279:1714–1718. doi: 10.1126/science.279.5357.1714. [DOI] [PubMed] [Google Scholar]
- Lang PJ, Davis M. Emotion, motivation, and the brain: reflex foundations in animal and human research. Prog Brain Res. 2006;156:3–29. doi: 10.1016/S0079-6123(06)56001-7. [DOI] [PubMed] [Google Scholar]
- Lee MG, Hassani OK, Alonso A, Jones BE. Cholinergic Basal Forebrain Neurons Burst with Theta during Waking and Paradoxical Sleep. J. Neurosci. 2005;25:4365–4369. doi: 10.1523/JNEUROSCI.0178-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee MG, Manns ID, Alonso A, Jones BE. Sleep-Wake Related Discharge Properties of Basal Forebrain Neurons Recorded with Micropipettes in Head-Fixed Rats. J Neurophysiol. 2004;92:1182–1198. doi: 10.1152/jn.01003.2003. [DOI] [PubMed] [Google Scholar]
- Lin SC, Gervasoni D, Nicolelis MA. Fast modulation of prefrontal cortex activity by basal forebrain noncholinergic neuronal ensembles. J Neurophysiol. 2006;96:3209–3219. doi: 10.1152/jn.00524.2006. [DOI] [PubMed] [Google Scholar]
- Maddux JM, Kerfoot EC, Chatterjee S, Holland PC. Dissociation of attention in learning and action: effects of lesions of the amygdala central nucleus, medial prefrontal cortex, and posterior parietal cortex. Behav Neurosci. 2007;121:63–79. doi: 10.1037/0735-7044.121.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto M, Hikosaka O. Lateral habenula as a source of negative reward signals in dopamine neurons. Nature. 2007 doi: 10.1038/nature05860. [DOI] [PubMed] [Google Scholar]
- McGaughy J, Dalley JW, Morrison CH, Everitt BJ, Robbins TW. Selective Behavioral and Neurochemical Effects of Cholinergic Lesions Produced by Intrabasalis Infusions of 192 IgG-Saporin on Attentional Performance in a Five-Choice Serial Reaction Time Task. J. Neurosci. 2002;22:1905–1913. doi: 10.1523/JNEUROSCI.22-05-01905.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muir J, Everitt B, Robbins T. AMPA-induced excitotoxic lesions of the basal forebrain: a significant role for the cortical cholinergic system in attentional function. J. Neurosci. 1994;14:2313–2326. doi: 10.1523/JNEUROSCI.14-04-02313.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolelis MA, Dimitrov D, Carmena JM, Crist R, Lehew G, Kralik JD, Wise SP. Chronic, multisite, multielectrode recordings in macaque monkeys. Proc Natl Acad Sci U S A. 2003;100:11041–11046. doi: 10.1073/pnas.1934665100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolelis MA, Ghazanfar AA, Faggin BM, Votaw S, Oliveira LM. Reconstructing the engram: simultaneous, multisite, many single neuron recordings. Neuron. 1997;18:529–537. doi: 10.1016/s0896-6273(00)80295-0. [DOI] [PubMed] [Google Scholar]
- Pang K, Williams MJ, Egeth H, Olton DS. Nucleus basalis magnocellularis and attention: effects of muscimol infusions. Behav Neurosci. 1993;107:1031–1038. doi: 10.1037//0735-7044.107.6.1031. [DOI] [PubMed] [Google Scholar]
- Parikh V, Kozak R, Martinez V, Sarter M. Prefrontal Acetylcholine Release Controls Cue Detection on Multiple Timescales. Neuron. 2007;56:141–154. doi: 10.1016/j.neuron.2007.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paton JJ, Belova MA, Morrison SE, Salzman CD. The primate amygdala represents the positive and negative value of visual stimuli during learning. Nature. 2006;439:865–870. doi: 10.1038/nature04490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 5th edition Academic Press; San Diego: 2005. [Google Scholar]
- Pessiglione M, Schmidt L, Draganski B, Kalisch R, Lau H, Dolan RJ, Frith CD. How the brain translates money into force: a neuroimaging study of subliminal motivation. Science. 2007;316:904–906. doi: 10.1126/science.1140459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson R, DeLong M. Context-dependent responses of primate nucleus basalis neurons in a go/no-go task. J. Neurosci. 1990;10:2528–2540. doi: 10.1523/JNEUROSCI.10-08-02528.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riekkinen P, Jr., Sirvio J, Riekkinen M, Riekkinen P. EEG changes induced by acute and chronic quisqualic or ibotenic acid nucleus basalis lesions are stabilized by tacridine. Brain Res. 1991;559:304–308. doi: 10.1016/0006-8993(91)90016-o. [DOI] [PubMed] [Google Scholar]
- Roesch MR, Olson CR. Neuronal activity related to reward value and motivation in primate frontal cortex. Science. 2004;304:307–310. doi: 10.1126/science.1093223. [DOI] [PubMed] [Google Scholar]
- Roitman MF, Wheeler RA, Carelli RM. Nucleus accumbens neurons are innately tuned for rewarding and aversive taste stimuli, encode their predictors, and are linked to motor output. Neuron. 2005;45:587–597. doi: 10.1016/j.neuron.2004.12.055. [DOI] [PubMed] [Google Scholar]
- Sambeth A, Maes JH, Van Luijtelaar G, Molenkamp IB, Jongsma ML, Van Rijn CM. Auditory event-related potentials in humans and rats: effects of task manipulation. Psychophysiology. 2003;40:60–68. doi: 10.1111/1469-8986.00007. [DOI] [PubMed] [Google Scholar]
- Sarter M, Bruno JP. The neglected constituent of the basal forebrain corticopetal projection system: GABAergic projections. European Journal of Neuroscience. 2002;15:1867–1873. doi: 10.1046/j.1460-9568.2002.02004.x. [DOI] [PubMed] [Google Scholar]
- Schultz W, Dayan P, Montague PR. A neural substrate of prediction and reward. Science. 1997;275:1593–1599. doi: 10.1126/science.275.5306.1593. [DOI] [PubMed] [Google Scholar]
- Semba K. Multiple output pathways of the basal forebrain: organization, chemical heterogeneity, and roles in vigilance. Behavioural Brain Research. 2000;115:117–141. doi: 10.1016/s0166-4328(00)00254-0. [DOI] [PubMed] [Google Scholar]
- Seymour B, O'Doherty JP, Koltzenburg M, Wiech K, Frackowiak R, Friston K, Dolan R. Opponent appetitive-aversive neural processes underlie predictive learning of pain relief. Nat Neurosci. 2005;8:1234–1240. doi: 10.1038/nn1527. [DOI] [PubMed] [Google Scholar]
- Voytko ML, Olton DS, Richardson RT, Gorman LK, Tobin JR, Price DL. Basal forebrain lesions in monkeys disrupt attention but not learning and memory. J Neurosci. 1994;14:167–186. doi: 10.1523/JNEUROSCI.14-01-00167.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward LM. Synchronous neural oscillations and cognitive processes. Trends Cogn Sci. 2003;7:553–559. doi: 10.1016/j.tics.2003.10.012. [DOI] [PubMed] [Google Scholar]
- Weinberger NM. The nucleus basalis and memory codes: auditory cortical plasticity and the induction of specific, associative behavioral memory. Neurobiol Learn Mem. 2003;80:268–284. doi: 10.1016/s1074-7427(03)00072-8. [DOI] [PubMed] [Google Scholar]
- Wenk GL. The nucleus basalis magnocellularis cholinergic system: one hundred years of progress. Neurobiol Learn Mem. 1997;67:85–95. doi: 10.1006/nlme.1996.3757. [DOI] [PubMed] [Google Scholar]
- Whalen P, Kapp B, Pascoe J. Neuronal activity within the nucleus basalis and conditioned neocortical electroencephalographic activation. J. Neurosci. 1994;14:1623–1633. doi: 10.1523/JNEUROSCI.14-03-01623.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiest MC, Bentley N, Nicolelis MA. Heterogeneous integration of bilateral whisker signals by neurons in primary somatosensory cortex of awake rats. J Neurophysiol. 2005;93:2966–2973. doi: 10.1152/jn.00556.2004. [DOI] [PubMed] [Google Scholar]
- Wilson FA, Rolls ET. Learning and memory is reflected in the responses of reinforcement-related neurons in the primate basal forebrain. J Neurosci. 1990;10:1254–1267. doi: 10.1523/JNEUROSCI.10-04-01254.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yacubian J, Glascher J, Schroeder K, Sommer T, Braus DF, Buchel C. Dissociable systems for gain- and loss-related value predictions and errors of prediction in the human brain. J Neurosci. 2006;26:9530–9537. doi: 10.1523/JNEUROSCI.2915-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaborszky L, Duque A. Sleep-wake mechanisms and basal forebrain circuitry. Front Biosci. 2003;8:d1146–1169. doi: 10.2741/1112. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.