
Tique
Ixodida
Cet article concerne un ordre d'acariens. Pour la plante appelée tique, voir Aextoxicon punctatum. Pour les volcans surnommés tiques, voir Volcanisme sur Vénus.
.jpg)
| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Chelicerata |
| Classe | Arachnida |
| Sous-classe | Acari |
| Super-ordre | Parasitiformes |
- Metastigmata Canestrini, 1891
Les Ixodida (du grec ancien ἰξώδης / ixôdês, « gluant »), appelées couramment tiques, sont un ordre d'arachnides acariens. Il est décrit par William Elford Leach en 1815. Cet ordre regroupe, en 2010, 896 espèces classées en trois familles[1] dont 41 en France, parmi lesquelles 4 très occasionnelles[pas clair][2]. Elles étaient autrefois appelées « Tiquet » ou « Ricinus ».
Les tiques sont des acariens ectoparasites, venimeux[3], de vertébrés (y compris des vertébrés à sang froid tels que les lézards, les serpents et les tortues).
Elles passent une partie de leur cycle au sol (éclosion, métamorphose et quête d'un hôte), et une autre partie (deux ou trois stades) ancrées sur la peau de mammifères (sauvages et d'élevage), d'oiseaux ou de reptiles, se nourrissant de leur sang grâce à un rostre. Leur digestion est extra-corporelle[4] ; elles peuvent à cette occasion transmettre à leurs hôtes de nombreux agents pathogènes connus (virus, bactéries, protozoaires, nématodes) responsables des maladies vectorielles à tiques, et parfois des neurotoxines (responsables de paralysie à tiques). L'homme peut développer des allergies à leur salive.
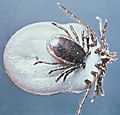
Ce sont surtout les femelles adultes nourries ou en train de se gorger de sang qui sont les plus repérables, car bien plus grosses que lors des autres stades de développement. À titre d'exemple, la pesée d'une femelle de la tique Hyalomma asiaticum avant et après son repas final indique un poids multiplié par 624[5],[6]. De tels repas permettent aux tiques de pondre de plusieurs dizaines à plusieurs centaines de milliers d'œufs (ce chiffre variant selon les espèces et selon les individus au sein de l'espèce).
Description
[modifier | modifier le code]




Les tiques sont des acariens métastigmates de grande taille (3 à 6 mm en moyenne, hors réplétion, et jusqu'à 30 mm pour certaines espèces tropicales), ce qui en fait les plus grands représentants de l'ordre des acariens. La forme, taille et couleur des tiques varient beaucoup selon l'espèce et son stade de développement[11] mais leur corps est toujours ovalaire et leur tête est prolongée d'un rostre équipé de deux chélicères.
Leur corps n'est pas segmenté en trois régions comme chez la plupart des arthropodes (on dit que leur métamérie est indistincte). On distingue simplement la partie antérieure (capitulum, « tête » en latin) de la partie postérieure (idiosome).
Capitulum
[modifier | modifier le code]Le capitulum est constitué d'une région basale (base capitulaire, ou basis capituli) qui porte trois pièces buccales (mais jamais d'yeux qui, chez les espèces qui en possèdent, sont disposés sur le scutum de l'idiosome) :
- un hypostome, dirigé vers l'avant, côté ventral, et garni de plusieurs files de dents qui aident la tique à s'ancrer solidement dans la peau de son hôte. La longueur et la forme du rostre sont utilisées en systématique ;
- une paire de chélicères, rétractiles dans une gaine protectrice, terminés par des dents ; ils sont dilacérateurs et coupent la peau, venant ainsi mécaniquement en aide à l'action chimique de la salive, pour perforer la peau. Ils se superposent à l'hypostome, en face dorsale, les deux pièces formant le rostre vulnérant. On distingue les tiques longirostres, dont le rostre est plus long que large, des brévirostres dont le rostre est aussi large ou plus large que long ;
- une paire de palpes (ou pédipalpes), au rôle sensoriel (ils ne sont pas vulnérants et ne pénètrent pas dans les tissus lors de la fixation de la tique, mais restent posés à la surface de la peau). Disposés latéralement au rostre, ils sont formés chacun de quatre articles de tailles et fonctions différentes :
- le premier, très court, articule le palpe sur la basis capituli,
- le deuxième et le troisième sont plus longs que le premier et pourvus de soies sensorielles,
- le quatrième est plus réduit, mieux visible du côté ventral, caché dans une fossette du troisième article, très richement pourvu de petites soies sensorielles.
Les tiques n'ont pas de canal alimentaire, ni de canal salivaire, comme la plupart des insectes hématophages (moustiques, punaises, etc.) ; l'apport de salive comme la succion du sang se font par l'espace ménagé entre hypostome et chélicères.
Le tégument des tiques est garni de pores, soies et divers autres organes sensoriels, visibles à la loupe binoculaire, dont les fonctions ne sont pas encore nettement établies parmi la détection de CO2, hormones, hygrométrie, température, vibrations… La détection du gaz carbonique et de l’acide butyrique dégagés par les animaux jouent un rôle dans le choix de l’hôte[12] et peut-être dans le choix du point de fixation de la tique sur son hôte. Au moins chez certaines espèces (ex. : Dermacentor andersoni[13], Dermacentor variabilis[13], Anocentor nitens[14]) les femelles émettent du 2,6-dichlorophénol (en), phéromone sexuelle très attractive pour les mâles de certaines espèces[13] et ne semblant pas générer de modification de comportement chez d'autres (ex : Boophilus microplus[15]).
Idiosome
[modifier | modifier le code]- Sa face dorsale porte une plaque sclérifiée dite « écusson » ou « scutum » couvrant environ la moitié du dos d'une tique à jeun, la totalité chez les mâles.
- Les 8 pattes s’insèrent en ligne sur la face ventrale. Elles comportent les articles classiques (coxa, trochanter, fémur, patelle, tibia, et tarse) et sont conclues par une ventouse (pulville) et deux griffes permettant à la tique de se déplacer sur presque tous les supports. La première paire de pattes porte au niveau du tarse un organe sensoriel olfactif important, l’organe de Haller, sensible[16],[17],[18] à la fois au degré d'hygrométrie, aux phéromones, au gaz carbonique, aux métabolites exhalés par les ruminants, à l'acide lactique, etc.
- Un pore génital s’ouvre sur la face ventrale, et nettement plus en arrière s'ouvre l’orifice anal. La position du « sillon anal » (antérieur ou postérieur à l’anus) différencie la famille des Ixodes des autres tiques dures. La forme et taille de l’écusson, la présence et position de stigmates, de soies sensorielles et d’ocelles… qui orientent la diagnose.
Symbiotes
[modifier | modifier le code]On connaît quelques symbiotes vivant dans la tique (ex. : Spiroplasma sp.) et des bactéries de l’environnement y sont aussi détectées (ex. : Mycobacterium sp.)[19], lesquels pourraient éventuellement interagir avec des micropathogènes véhiculés par la tique.
Systématique
[modifier | modifier le code]Les Ixodida comptent trois familles actuelles :
- Ixodidae C. L. Koch, 1844 (ou tiques dures) ; environ 670 espèces connues, possédant toutes des zones de tégument chitinisé dur ;
- Argasidae C. L. Koch, 1844 (ou tiques molles) ; environ 180 espèces aux téguments non sclérifiés ;
- Nuttalliellidae Schulze, 1935, une famille intermédiaire entre les deux précédentes, monogénérique et monospécifique.
Parmi les familles éteintes :
- †Deinocrotonidae Peñalver et al., 2017[20].
La phylogénie place les tiques comme le groupe frère des Holothyrida.
- Les Ixodidae ou tiques dures
Elles sont les mieux connues et étudiées car rassemblant la plupart des tiques adaptées aux animaux domestiques et/ou mordant l'Homme. Elles causent des maladies et l'affaiblissement des animaux qu'elles infestent, et sont ainsi sources d'importantes pertes économiques pour certains éleveurs, de même pour les systèmes de santé publique et de sécurité sociale, notamment en raison de la maladie de Lyme, dont certains ixodes sont vecteurs[21] en pleine expansion[22].
-
 Comparaison avec une allumette.
Comparaison avec une allumette. -
 Une femelle de l'espèce Ixodes hexagonus
Une femelle de l'espèce Ixodes hexagonus -
 Femelle de 4 mm environ (Iéna, Allemagne)
Femelle de 4 mm environ (Iéna, Allemagne) -
 Mâle et femelle d’Ixodes ricinus copulant. Le mâle est le plus petit
Mâle et femelle d’Ixodes ricinus copulant. Le mâle est le plus petit -
 Ixodes scapularis, un des vecteurs de la maladie de Lyme
Ixodes scapularis, un des vecteurs de la maladie de Lyme
.jpg)
.jpg)



Les tiques de la famille des Ixodidae sont de taille très variable, avec un capitulum antérieur bien visible et un scutum dorsal dont la présence est constante. Elles vivent dans des biotopes très variés, soit dans des abris fermés correspondant aux abris de leurs hôtes, soit à ciel ouvert, dans la végétation basse, où elles pondent, muent, et guettent, au sommet des herbes, leur hôte pour prendre leur repas de sang. Contrairement à ce qui a longtemps été colporté, les tiques ne tombent pas des arbres, la proximité avec le sol, pour des raisons de réhydratation, étant essentielle. En général, dans la végétation, la dispersion des individus est la règle, mais des regroupements sur une même herbe sont possibles, par l'effet des phéromones (médiateurs chimiques servant à la communication entre tiques) ; des concentrations spatiales plus larges s'observent aussi, notamment au niveau de biotopes favorables, par exemple autour des points d'eau, ou de zones d'ombre, ou de regroupement ou passage d'animaux… Les tiques se détachent de leur hôte, gorgées de sang, après un repas qui dure rarement plus d'une semaine pour le stade adulte femelle, moins pour les autres stades.
La tique la plus commune en Europe, Ixodes ricinus, appartient à cette famille.
En Amérique du Nord, la souris à pattes blanches (Peromyscus leucopus), est l'hôte principal d'Ixodes scapularis (tiques à pattes noires) aux stades larvaire et nymphal, vectrice de la maladie de Lyme. Les adultes sont également communs chez le cerf de Virginie (Odocoileus virginianus), au printemps et à l'automne principalement, mais cette tique a été trouvée chez plus de 50 autres espèces de mammifères et chez plus de 55 espèces d'oiseaux.
La tique sénégalaise Amblyomma variegatum présente en Afrique et dans les Caraïbes sur, entre autres, les animaux d'élevage, appartient à cette famille. Elle pourrait peut-être paradoxalement être disséminée par le héron garde-bœuf (Bubulcus ibis)[23], qui se nourrit des parasites de grands mammifères.
- Les Argasidae ou tiques molles
Ce sont les plus grosses. Leur rostre situé sur le ventre est invisible en vue dorsale. Elles vivent généralement près de leur hôte dans les crevasses, terriers, nids ou habitations et viennent se nourrir plusieurs fois sur leur hôte lorsque celui-ci est immobile. Elles peuvent jeûner jusqu'à 5 ans.
En Europe, elles sont surtout présentes dans le pourtour méditerranéen.
- Les Nuttalliellidae
Peu connues, elles partagent des caractéristiques des deux autres familles. Une seule espèce a été décrite, Nuttalliella namaqua.
Reproduction et cycle de développement
[modifier | modifier le code]Seules les tiques adultes s'accouplent, au sol avant la quête de l’hôte ou sur l’hôte lui-même. Après l'accouplement, le mâle meurt ; la femelle meurt après la ponte[24].
Toutes les espèces connues de tiques se développent en passant par quatre stades évolutifs distincts :
- L'œuf : certaines espèces pondent un très grand nombre d'œufs : on en a par exemple compté 23 891 dans une seule ponte d'Amblyomma nuttalli, une tique africaine qui parasite les tortues[25] ;
- La larve (qui n'a que 3 paires de pattes, griffues) ;
- La nymphe (qui est octopode comme l'adulte mais sans orifice génital) ;
- L'adulte : à ce stade le dimorphisme sexuel est généralement net (écusson chitinisé) plus développé en face dorsale chez le mâle des Ixodidae que chez la femelle, dont le tégument doit rester souple pour se distendre pour une prise de sang maximale afin d'élaborer ses œufs.
Dans la plupart des cas, durant son cycle de vie, une tique change d'hôte à chacun de ces stades :
- le cycle est dit triphasique quant au sortir de l'œuf, la larve se fixe sur un premier hôte qu'elle quitte après y avoir fait son premier repas, c'est-à-dire effectué sa première phase parasitaire. Elle mue alors au sol, puis doit trouver un nouvel hôte pour son repas nymphal, qui est la deuxième phase parasitaire, après quoi elle se laissera à nouveau tomber au sol pour effectuer la mue de transformation en adulte. Devenue adulte, elle recherche un troisième hôte pour effectuer le dernier repas ou troisième phase parasitaire (chez ces tiques, les adultes sont toujours plus porteurs de pathogènes transmissibles que la nymphe et la larve, chaque repas étant un risque de contracter les germes). Exemple : Rhipicephalus sanguineus a un cycle triphasique monotrope (c'est-à-dire effectué sur la même espèce-hôte aux trois stades) ;
- un petit nombre d'espèces de tiques ne changent pas d'hôte à chaque stade :
- le cycle est dit diphasique quand il ne comporte que deux phases parasitaires : la larve effectue son premier repas sur un hôte, puis mue en nymphe et effectue son second repas sur le même hôte avant de se détacher pour se transformer en une tique adulte qui effectuera sa deuxième phase parasitaire et son troisième repas sur un deuxième hôte,
- le cycle est dit monophasique quand la tique effectue tout son cycle de développement (trois repas et deux mues) sur un même individu-hôte. Seule la larve à jeun effectue une quête d'hôte. Exemple : Rhipicephalus microplus (en).
Stratégie de quête d'hôte et alimentation
[modifier | modifier le code]Pendant les phases de « quête », la tique attend un hôte pour s'y accrocher. La quête se fait en milieu extérieur ouvert (sur les brins d'herbe, de graminées, de fougères, etc.) pour les tiques exophiles ; elle se fait au sol (à même la terre ou sur les brindilles des nids et terriers ou les crevasses des grottes) pour les tiques endophiles. Parmi les exophiles, différentes stratégies s'observent :
- dispersion des tiques sur un large territoire, sur tous types de végétation (surtout dans les zones à conditions climatiques peu sévères) ;
- distributions spatiales et temporelles ciblant des lieux où les chances de rencontre avec l'hôte sont plus favorables : zone d'ombres et de nourrissage en zone aride pour les tiques infestant les mammifères, etc. Par exemple, les larves de Rhipicephalus appendiculatus ou Rhipicephalus zambeziensis en saison sèche seront surtout trouvées dans la végétation entourant les trous d'eau permanents ou sur des herbes dominées par des Acacias, c'est-à-dire là où des ongulés ou d'autres mammifères viendront nécessairement se nourrir et/ou s'abriter du soleil ;
- des larves semblent aussi pouvoir se laisser transporter en aval par l'eau lors de crues. Elles pourront sur une berge plus éloignée tenter de trouver un autre hôte. En zone aride, les cours d'eau attirent inévitablement de nombreux animaux qui y viennent boire. Ces animaux pourront à leur tour véhiculer la tique, parfois sur de grandes distances, ce qui permet l'entretien de vastes métapopulations, génétiquement riches et donc très adaptables aux modifications de leur environnement (et aux pesticides antiparasitaires le cas échéant) ;
- certaines tiques se déplacent remarquablement bien sur leur hôte (même animaux lisses à écailles tels que certains serpents, orvets, lézards) ou percent une peau épaisse (de rhinocéros ou de tortue), sans que l'hôte les sente ni cherche à se débarrasser d'elles ;
- la salive de tique contient un cocktail de molécules[26],[27],[28],[29],[30],[31],[32] : outre des enzymes digestifs cytolytique[33], cette salive contient notamment des molécules anesthésiantes, qui rendent la morsure indolore et la tique moins détectable par l'hôte. D'autres molécules sont anticoagulantes[33] et vasodilatatrices[33], et d'autres encore inhibent la réponse immunitaire de l'hôte[34],[35]. Cette salive est aussi un venin ; elle contient des neurotoxines et enzymes proches de composés présents dans les venins d’araignée et de scorpion[3]. (rem : avec sa salive, parmi divers pathogènes, la tique peut notamment parfois - de plus - inoculer un pathogène immunodépresseur : Anaplasma phagocytophilum) ; « Une protéine de salive de tique, Salp15, a été montrée comme essentielle pour la transmission de B. burgdorferi. Cette protéine inhibe l’activation des lymphocytes et la maturation des cellules dendritiques et se fixe à une protéine de surface majeure de B. Burgdorferi, OspC (Outer surface protein C), pour faciliter la transmission bactérienne et protéger les bactéries contre les anticorps de l’hôte »[35] ; des extraits de salive de tique inhibent ou modulent la réponse immune[36] et inflammatoire de la peau (des kératinocytes), avec des effets anti-sérotonine[37], anti-histamine[37], anti-anaphylactique[38], antialarmine[35],[39] ;
- libres ou attachées, elles sont très résistantes à l'écrasement et montrent une étonnante capacité à échapper aux différentes méthodes de nettoyage de leur hôte (par léchage, grattage, bains de boue ou de poussière, autoépouillage). L'épouillage réciproque, véritable rite social développé par les singes semble une des méthodes les plus efficaces, bien que longue.
Ces différentes stratégies peuvent être combinées, avec des variations saisonnières (ex. : concentration autour de l'eau douce en saison sèche et chaude). Les larves étant mobiles, mais sur d'assez courtes distances, elles peuvent en cas d'insuccès tenter de se déplacer et chercher un lieu plus favorable (ce comportement semble plus fréquent quand il fait plus chaud, chez I. ricinus).
Ancrage de la tique et repas sur l'hôte
[modifier | modifier le code]

Après avoir trouvé une proie et s'y être accrochée, la tique chemine lentement sur la peau (de quelques minutes à plusieurs heures parfois) pour trouver un emplacement qui lui convient. De fines griffes lui permettent de se stabiliser sur l'épiderme (ces griffes sont plus puissantes chez la larve qui a besoin de s'ancrer pour pouvoir percer la peau). La tique coupe la peau grâce à des chélicères extériorisables (cachées au repos dans une gaine protectrice) qu'elle enfonce peu à peu ainsi que l'hypostome, aidé par la sécrétion d'enzymes salivaires (protéases) qui provoquent une cytolyse. Au bout de son rostre se forme alors une poche ou chambre de cytolyse. Cette opération, sous l'effet de substances salivaires anesthésiantes, se fait sans douleur pour l'hôte. La tique en quelques heures a ainsi enfoncé tout son rostre. Elle parfait son ancrage par la sécrétion d'une substance, sorte de colle biologique dite « cément » (ou « manchon hyalin ») ; cette colle la fixe très fortement au derme. Ainsi fixée, elle peut alors, pendant toute la durée de son repas, alternativement[2] aspirer le sang et réinjecter de la salive de manière à agrandir la poche ainsi creusée sous la peau jusqu'à ce que cette poche atteigne un ou plusieurs microcapillaires sanguins, qui crèveront et l'alimenteront directement en sang. Durant ce temps, la tique injecte un cocktail de molécules qui affaiblissent localement l'immunité de l'hôte et insensibilisent le système nerveux (ce qui ne fonctionne plus chez des organismes dont le système immunitaire a été sensibilisé, rendu allergique en quelque sorte, à ces molécules). La tique dispose aussi de moyens de détecter et tuer une partie des bactéries pathogènes qu'elle ingère lors de son repas, via des lysozymes présents dans sa salive, notamment[40].
Plus le rostre est long, mieux la tique est fixée. Ainsi des tiques brévirostres telles que Rhipicephalus ou Dermacentor s'ôtent facilement de la peau, n'y laissant que leur manchon hyalin, alors que des tiques longirostres, telles qu'Ixodes et Amblyomma, sont si bien fixées qu'une traction directe leur arrache souvent le rostre, lequel peut causer un abcès ou une infection.
Une première publication a décrit en 2005 une neurohormone peptidergique, isolée chez Ixodes ricinus et Rhipicephalus microplus[41]. Ce peptide (dit Ixori-PVK, PALIPFPRV-NH2) est proche (homologie de séquence) des peptides periviscerokinin/CAP2b des insectes, impliquées dans la régulation du bilan hydrique. Elle pourrait donc jouer un rôle important dans le processus diurétique lié au repas de sang relativement « liquide »[41].
Habitats, densités et aires de répartition
[modifier | modifier le code]Habitats
[modifier | modifier le code]Les habitats varient selon les espèces. Certaines sont dépendantes de l'humidité ; elles sont inféodées aux milieux forestiers, péri-forestiers ou boisés, ou confinées dans des abris de taille réduite permettant le maintien de l'humidité : nids, terriers, grottes, étables… (voir la faune des tiques de France[2]). D'autres espèces sont adaptées aux climats secs, voire désertiques.
Selon leurs exigences en matière d'habitats (auxquels elles sont plus ou moins inféodées), on classe les tiques en :
- espèces hygrophiles (des milieux humides) ;
- espèces xérophiles (des milieux secs)[42].
Elles sont aussi classées en :
- espèces exophiles : c'est-à-dire non inféodées à un biotope de petites dimensions. Ce sont des tiques qui sont dispersées dans la végétation de lisière, les fourrés ou les bordures de pâturages. Elles chassent à l’affût sur la végétation basse soit des petits animaux, surtout aux stades larvaires et nymphaux, soit de grands animaux, carnivores, ongulés… lorsque la tique est adulte ;
- espèces pholéophiles ou endophiles ou nidicoles (vivant dans les nids) : inféodées (à un seul ou à plusieurs stades de leur développement, y compris mues, pontes et phases de repos) à des habitats spécialisés ou à des conditions thermohygrométriques particulières (terrier de rongeur en général), caractérisés par le noir et un microclimat très stable (avec humidité élevée). En zone froide et en hiver, la température d'un animal hibernant suffit à protéger les larves, nymphes ou œufs de certaines tiques du gel ou de l'aridité du climat extérieur. Certaines espèces accomplissent la totalité de leur cycle dans un terrier, et très rarement dans nos habitations (plus fréquemment dans des chenils, étables…) en se nourrissant alors sur des animaux domestiques (Rhipicephalus sanguineus est un exemple de tique devenue « domestique » ou endophile). Leurs "hôtes" sont plutôt des reptiles, micromammifères ou petits mammifères (lapins, lièvres, hérissons) ou des oiseaux (qui contribuent alors à leur large dispersion)[43],[44],[45] ;
- espèces mixtes : leurs larves et nymphes sont généralement pholéophiles alors que les adultes seront exophiles.
Les genres plus primitifs de tiques (Ixodes et Haemophilus) sont plutôt forestiers et les genres plus évolués (Dermacentor) « privilégient des formations végétales ouvertes : steppes et prairies »[46].
La plus grande variété d'espèces se rencontre en zones chaudes. Les microhabitats qu'elles occupent varient selon l'espèce et le stade de développement. Les larves et nymphes ont besoin d'une humidité relative plus élevée (50 à 80 % du seuil de saturation selon les espèces). Les adultes de certaines espèces supportent plusieurs heures de climat aride, en se réhydratant la nuit et le matin avec la rosée.
Pour de très nombreuses espèces de tiques (en forêt tropicale notamment), les hôtes sont mal connus et les exigences des œufs, des larves et des nymphes plus encore.
Aires de répartition et densités de tiques
[modifier | modifier le code]Des tiques peuvent vivre ou survivre presque partout sur les terres émergées, sauf en zone trop aride et chaude ou trop froide.
Dans de vastes parties du monde, l'aire et les densités de certaines espèces (et par suite de la maladie de Lyme) semblent rapidement s'étendre depuis deux ou trois décennies et se déplacer avec le réchauffement climatique et pour d'autres raisons (changement de comportement des tiques). Ainsi, selon une étude récente parue dans le Journal of Medical Entomology (2016), deux espèces de tiques (qui sont les principaux vecteurs de la maladie aux États-Unis : Ixodes scapularis et I. pacificus) ont depuis 20 ans fortement étendu leur aire de répartition : L’une ou l’autre de ces deux tiques est maintenant trouvée dans la moitié de tous les comtés des États-Unis. Les populations de tiques à pattes noires (I scapularis) ont géographiquement doublé en moins de 20 ans pour être aujourd’hui signalées dans 45,7 % des comtés américains (dans 37 États à travers l'est des États-Unis), et en hausse de 30 % en 1998[47]. La tique à pattes noires de l’ouest est plus rare, à ce jour uniquement détectée dans six États, mais elle a aussi montré des augmentations démographiques et géographiques, bien que plus modestes (extension géographique : + 3,4 % à 3,6 %)[47]. Les écoépidémiologistes sont préoccupés par le fait que c’est au nord-est que les densités de tiques sont les plus élevées, précisément là où la maladie de Lyme est devenue la plus courante[47].
Une étude de 2015 apporte un début d'explication à ces phénomènes : les populations de tiques à pattes noires ont un comportement qui diffère dans le nord et le sud du pays : Au nord, les nymphes en quête de nourriture sont plus actives et « audacieuses » ; ainsi, dans le Wisconsin et dans l'état de Rhode Island, elles se montrent 20 fois plus susceptibles d'émerger de la litière de feuilles et de se mettre sur le chemin des humains, par rapport à des nymphes prélevées dans le Tennessee et en Floride. Les auteurs craignent que les tiques du nord se développent vers le sud[47].
En 2017, dans le New Jersey, les autorités américaines s'inquiètent de voir se développer une espèce nouvelle de tiques d'Asie particulièrement invasive et dangereuse[48].
Dynamique des populations
[modifier | modifier le code]L'importance des populations et de leur dynamique est difficile à évaluer précisément.
Des techniques de capture-recapture ont récemment (2016-2017) été testées[49]
Peu des œufs pondus donneront une tique adulte, mais chaque femelle pond des milliers d'œufs.
Les œufs, larves ou adultes de nombreuses espèces de tiques meurent quand il fait trop froid ou trop sec. Si l'humidité du sol suffit généralement à protéger les larves de la dessiccation, une forte humidité au sol et/ou une hygrométrie élevée favorisent plusieurs espèces de nématodes et de champignons entomopathogènes qui peuvent infecter et tuer les tiques, soit directement soit via des bactéries symbiotiques.
Les larves, nymphes et adultes peuvent aussi directement être mangées au sol par des oiseaux, reptiles et autres animaux insectivores (une poule peut ainsi manger 200 tiques par heure dans une zone fructueuse[50]), voire sur leur hôte (par exemple par le héron garde-bœufs ou lors de comportements d'épouillage), mais les animaux qui mangent des tiques servent aussi d'hôtes à diverses espèces de tiques, dont celles qu'ils mangent. Cependant des oiseaux (dont prédateurs de tiques tel que de héron garde-bœufs, Bubulcus ibis) contribuent aussi à disperser des larves de tiques lors de leurs migrations[51].
La prolificité des tiques, et leur résistance quand leurs conditions de vies sont bonnes, laissent penser qu'elles pourraient s'adapter aux acaricides et antiparasitaires, voire à certains moyens de lutte biologique.
Quelques recherches portent depuis peu sur diverses stratégies de lutte biologique contre ces hématophages[52].
Prédateurs
[modifier | modifier le code]Les tiques (œufs, larves, nymphes et imago) semblent dans la nature surtout contrôlés par quelques animaux insectivores et par plusieurs catégories de microorganismes parasites et « entomopathogènes » encore peu étudiés et méconnus[53]. Parmi les prédateurs connus des tiques, figurent :
- des reptiles ; certains lézards ou des orvets mangent les tiques et leurs larves ; certaines espèces de lézards très parasitées dans la nature consomment une grande partie des tiques qui tenteront de s'accrocher sur elles (83 à 96 % pour certains lézards ; ces derniers peuvent alors — en tant qu'espèce — détruire des milliers de tiques par hectare, en jouant aussi un rôle de piège écologique pour les larves qui auront tenté de les parasiter) alors que d'autres espèces de reptiles semblent ne pas réagir (favorisant alors les tiques, mais pas les borrélies, qui ne se reproduisent pas chez cet hôte à sang froid)[54] ;
- des mammifères insectivores ; en Europe ce sont principalement les musaraignes, les taupes et le hérisson, ce dernier ne pouvant toutefois pas se débarrasser de la plupart des tiques qui se fixent sur lui. En Amérique du Nord, les opossums et les écureuils se montrent très souvent parasités, mais une étude récente a montré qu'ils mangent 83 à 93 % des I. scapularis qui tentent de se fixer sur eux, contribuant à réguler les populations de ces tiques et des pathogènes qu'elles véhiculent[54] ;
- des oiseaux ; les espèces comme le piquebœuf à bec rouge (Buphagus erythrorhynchus) ou le piquebœuf à bec jaune (Buphagus africanus) sont depuis longtemps connus comme mangeurs de parasites, pouvant absorber jusqu'à plus de 12 000 larves de tiques ou une centaine de femelles engorgées par jour (M.Stutterheim et al. 1998). Le héron garde-bœufs (Bubulcus ibis), parfois dit « tick bird », mange aussi des tiques[55]. Certains passereaux (dont les étourneaux) se nourrissent aussi de tiques au sol et parfois même directement sur des bovins qu'ils déparasitent. Au Kenya des vétérinaires ont aussi débarrassé des vaches et chèvres africaines de leurs tiques en les mettant en contact avec des poules dans de petits enclos ; une seule poule peut ainsi manger jusqu'à 200 tiques en trois heures. La pintade de Numidie (Numida meleagris) ne semble par contre ne manger que les tiques adultes gorgées de sang, alors que les nymphes sont bien plus nombreuses et responsables de la plupart des cas d'infections[53] ;
- des insectes comme les larves de Chrysope, des coléoptères dont la plupart des carabes autochtones selon F. Gigon (ex. : Carabus problematicus et C. glabratus) et certaines espèces de fourmis, et sans doute d'autres espèces non encore identifiées. Les coléoptères semblent préférer les tiques déjà gorgées de sang, peut être parce qu'ils les repèrent plus facilement que les larves ou nymphes souvent immobiles durant leur quête d'un hôte de passage[56].
Plusieurs guêpes parasitoïdes parasitent les tiques. On voudrait les utiliser pour la lutte biologique contre les tiques, mais on ne sait pas encore les élever en masse. Ixodiphagus hookeri (Encyrtidae) antérieurement nommée Hunterellus hookeri ou Ixodiphagus caucurtei est l'un des principaux parasites de tiques du genre Ixodes ricinus[52]. On connaît au moins huit guêpes parasitoïdes de tiques. Ixodiphagus hookeri est assez ubiquiste et présente en France, mais aussi en Allemagne, au Royaume-Uni, en Moldavie ou au Portugal (selon les données Zipcodezoo.com consultée le 2009-09-28) [771, 918][source insuffisante] (elle parasite de 7 à 20 % des I. ricinus en France selon JM Doby (1993)[réf. nécessaire], s'attaquant à un large panel d'espèces (Dermacentor, Haemaphysalis, Hyalomma, Ixodes, Rhipicephalus, toujours aux stases infra-imaginales)[57]…
Ixodiphagus hookeri, qui vit moins de trois jours au stade adulte, pond un ou plusieurs œufs dans des larves ou des nymphes de tiques non gorgées, ces œufs ne se développant que stimulés par le repas de sang de leur hôte, ce qui est rendu possible par un virus symbiotique qui permet à l’œuf de ne pas être détecté par le système immunitaire de la tique, puis à la larve sortie de cet œuf de pouvoir dévorer la tique de l'intérieur[58] ; - des araignées ;
- d'autres acariens, dits entomophages ;
- des amphibiens, durant leur stade terrestre de vie ;
- des microorganismes :
- champignons parasites entomopathogènes, dont Metarhizium anisopliae, Beauveria bassiana[59], Paecilomyces fumosoroseus qui s'avèrent en laboratoire rapidement mortels pour les tiques Ixodes ricinus, en 5 à 25 jours… En lutte biologique, des blastospores pourraient même être encore plus efficaces que des conidiospores[52],
- nématodes entomopathogènes, dont plusieurs sous-espèces de Steinernema et d'Heterorhabditis qui sont capables d'inoculer leurs bactéries symbiotes (Xenorhabdus, Photorhabdus) dans les tiques qu'ils parasitent, qui en meurent rapidement. Ces bactéries liquéfient en effet l'intérieur de la tique (lyse) que les nématodes peuvent ensuite digérer[60],[61]. Ces nématodes ne se reproduisent pas dans la tique, mais ensuite dans l'eau interstitielle du sol ou dans le sol humide. Des forêts trop drainées, trop sèches et trop éclairées ne leur conviennent pas (comme presque tous les nématodes, ces espèces sont lucifuges).
In vitro, 10 à 40 % des tiques femelles non alimentées et contaminées meurent ; S. carpocapsae semblant être le nématode le plus souvent mortel pour ces tiques, d'autant plus que sa densité dans le milieu est élevée (deux fois plus de mortalité avec un « ensemencement » de 600 nématodes/cm2 de substrat que pour 300 nématodes/cm2)[52], - bactéries pathogènes pour les tiques : par exemple, on a découvert par hasard dans une colonie de tiques (Ornithodoros moubata) élevée en laboratoire une bactérie à pigment jaune, Gram-négatif (Chryseobacterium indologenes) qui se reproduit dans l'intestin des tiques, en tuant une grande partie des tiques molles qu'elle infecte (100 % de mortalité si plus de 106 bactéries/ml), mais non les tiques dures qui s'en débarrassent en peu de temps[62].
Facteurs de pullulation des tiques
[modifier | modifier le code]L'aire de distribution des tiques (sauf peut-être en zone très aride) s'étend nettement, notamment en altitude. Ceci va probablement s'accompagner d'une augmentation des régions touchées par les maladies transmises par les tiques.
La recherche[63] évoque plusieurs causes, complémentaires et aggravantes, de pullulation :
- les modifications climatiques, favorisant en zone froide et tempérée la survie hivernale des tiques, et favorisant leur remontée en altitude ; le facteur climatique pour certaines espèces semble être un des bons signes annonçant des pullulations ou changements d'aire de répartition[64] ;
- des modifications environnementales profondes, d'origine anthropique, incluant :
- une raréfaction des ennemis des tiques en liaison avec une large contamination de l'environnement par les insecticides, qui pourrait expliquer un recul important des insectes et acariens parasites-prédateurs, notamment les guêpes parasitoïdes, ainsi que par les fongicides, avec la régression des nombreuses espèces de champignons entomopathogènes, avec aussi un recul des nématodes parasites des tiques,
- des modifications paysagères : en Amérique du Nord, les tiques seraient plus nombreuses dans les forêts plus fragmentées, peut-être en raison de l'augmentation des cultures en lisière. Les chênaies et les années de bonnes glandaies favorisent des pullulations de tiques (en favorisant des rassemblements d'animaux qui leur fournissent des repas de sang et des moyens de transport)[65]. Les petits rongeurs[66] et notamment la souris à pattes blanches sont ainsi fortement favorisée et cet animal est connu pour jouer un rôle majeur dans l'écoépidémiologie de la maladie en Amérique du Nord[67]. Les glandaies comptent en Amérique du Nord parmi les meilleurs prédicteurs d'augmentation de risque pour la maladie de Lyme[68],
- la densification des hôtes des tiques artificiellement alimentés : rongeurs, sangliers et cervidés notamment très favorisés par l'agrainage, mais aussi par l'étendue croissante des cultures céréalières en lisière forestière, et la perte de milliards de grains de céréales le long des axes de transport (route, voies ferrées) après les moissons, etc. (quantifiée sur certains axes aux États-Unis, car attirant des sangliers qui causent des accidents),
- la densification des hôtes liée à la disparition de leurs prédateurs, pourchassés ou décimés par les polluants. Les zones de pullulation de tiques correspondent souvent à des zones où les grands prédateurs ont disparu et où le grand gibier est (sur)alimenté.
Relations hôte-espèce
[modifier | modifier le code]Des tiques semblent pouvoir s'attaquer à presque tous les vertébrés terrestres (même munis d'écailles serrées comme les reptiles), mais la plupart des espèces sont spécialisées pour un groupe d'hôte, tout en pouvant, comme le font certaines tiques d'oiseaux, accidentellement mordre l'Homme quand elles sont en contact avec lui. Les mécanismes qui font qu'une tique soit capable de sélectionner sa proie sont encore mal connus, mais semblent faire intervenir l'odorat et un tropisme pour des conditions microclimatiques qui sont celles de son hôte.
- Espèces ubiquistes : Ce sont souvent des tiques exophiles. À tout ou partie de leur stade de développement, elles peuvent se nourrir sur une grande variété d'espèces (En Europe, c'est par exemple le cas des immatures d'Ixodes ricinus).
- Espèces monotropes : Ce sont les espèces qui - à leurs trois stades de développement - parasitent toujours la même espèce d’hôte (ex. : Rhipicephalus sanguineus).
- Espèces ditropes : Ce sont les tiques dont la larve et la nymphe se nourrissent sur un certain type d'hôte (micromammifères, les oiseaux et les reptiles en général) alors que les adultes ne parasitent que des grands mammifères (ex. : Dermacentor sp).
- Espèces télotropes : Ce sont les tiques dont les larves et les nymphes sont très ubiquistes alors que les adultes sont très sélectifs. (ex. : Ixodes ricinus).

L’apparition des tiques, puis leur diversification en plus de 900 espèces aujourd’hui, ont été grandement conditionnées par des associations symbiotiques avec des bactéries intracellulaires[69]. L'alimentation des tiques, uniquement composée de sang, pose un problème de carence en certains nutriments indispensables à leur cycle de vie, comme les vitamines B (B2, B8[a], B9). Ces vitamines sont peu présentes dans le sang des vertébrés et les tiques, tout comme les autres hématophages strictes, ne peuvent pas les synthétiser elles-mêmes[69]. Une étude de 2018 montre que les tiques obtiennent ces vitamines B grâce à une association unique avec des bactéries symbiotiques, appartenant au genre Francisella (en)[71],[72] et présentes chez au moins 20 % des espèces de tiques[73],[74]. Pour les autres espèces de tique, ce sont d'autres bactéries symbiotiques, principalement appartenant au genre Coxiella, qui assurent cet approvisionnement en vitamines B[73],[74]. Ces bactéries symbiotiques sont massivement présentes dans les tubes de Malpighi et ovaires des tiques, et se transmettent d'une génération à l'autre par la voie transovarienne[75],[72],[76]. Privées de ces bactéries symbiotiques, les tiques ne peuvent ni atteindre l'âge adulte, ni se reproduire, et seul un complément artificiel en vitamines B permet de restaurer une croissance normale[77],[72],[78]. La capacité de ces bactéries à synthétiser des vitamines B dérive d'un ancêtre pathogène dont le génome s'est progressivement dégradé, avec notamment la perte de gènes de virulence[72]. En raison de cette proximité génétique, ces bactéries symbiotiques sont souvent confondues avec des agents pathogènes, ce qui conduit à une surestimation des risques infectieux associés aux tiques[79],[80].
Paléontologie
[modifier | modifier le code]
Les tiques existent depuis plus de 140 millions d'années comme en témoignent des fossiles de diverses espèces trouvés dans plusieurs régions du monde. Elles seraient apparues au Crétacé (de −146 à −65 millions d'années) et auraient le plus évolué et se seraient le plus dispersées durant l'ère tertiaire (à partir de −65 millions d'années)[81]. La famille la plus ancienne connue par les fossiles est celle des Argasidae qui parasitaient les oiseaux du crétacé (trouvées dans des morceaux d’ambre du New Jersey). D’autres fossiles de tiques ont été trouvés dans de l’ambre de la mer Baltique ou de la république dominicaine. En 2017, la tique Deinocroton draculi est trouvée parmi des plumes de dinosaures, préservée dans de l'ambre de Birmanie datant de 99 millions d'années[20].
Des études datant de 2011 tendent à prouver que les premières tiques seraient originaires du bassin de Karoo dans le Gondwana à la fin du Permien supérieur, soit il y a environ 250 millions d'années[82].
Tiques et maladies
[modifier | modifier le code]Les tiques seraient dans le monde - si on considère la totalité de leurs populations - les vecteurs du plus grand nombre de micro-organismes différents[83]. Ce sont des agents vecteurs de nombreuses pathologies humaines et animales.
Elles véhiculent notamment des maladies émergentes, réémergentes humaines ou animales[84] ou qui pourraient le devenir :
- borréliose, ou maladie de Lyme, due à Borrelia burgdorferi, B. garinii, B. afzelii, etc. ;
- peste porcine africaine (en zone tropicale), due à un pestivirus ;
- fièvre du Nil occidental (en zone tropicale, et peut-être tempérée) ;
- rickettsiose (maladies sévères graves, dont la fièvre boutonneuse méditerranéenne et le typhus[85]), dues à des rickettsies ;
- anaplasmoses (dont l'anaplasmose bovine), dues aux Anaplasma ;
- coxiellose due à Coxiella burnetii ;
- babesiose, due à Babesia ;
- bartonellose, due à Bartonella, via au moins la tique Ixodes ricinus (transmission expérimentale effective pour Bartonella henselae[86], avec de forts arguments épidémiologiques en faveur d'une transmission vectorielle naturelle[87],[88] ;
- franciselloses due à Francisella (dont la tularémie) ;
- leishmaniose, due à Leishmania infantum, via la tique Rhipicephalus sanguineus dont les glandes salivaires abritent naturellement ce pathogène[89], avec passage transovarien (« de mère en fille ») démontré chez cette même tique[90], sans qu'on ait encore mesuré l'effectivité de la transmission dans la nature[91] ;
- diverses viroses, dues à des virus (dont des arbovirus) ;
- ehrlichiose ou cowdriose, due à la bactérie Ehrlichia ruminantium[92].
Les morsures de tiques peuvent parfois être causes d'urticaire ou de choc allergique (Choc anaphylactique[93]). Ces réactions sont plus ou moins fréquentes selon les espèces : de nombreux cas sont documentés pour Argas reflexus[94], Ixodes ricinus[95], Ixodes holocyclus[96] et Ixodes pacificus[97],[98], mais l'allergie semble plus rare pour les Rhipicephalus[98].
L'épidémiologie et l'écoépidémiologie de ces maladies pourraient être modifiées par des bactéries symbiotiques[99].
Des tiques, dures et molles, pourraient aussi jouer un rôle important dans la conservation hivernale de virus réputés transmis par d'autres espèces ; on a récemment expérimentalement montré[100] (étude par PCR en temps réel) chez plusieurs espèces de tiques (Ixodes ricinus, Ixodes hexagonus, Dermacentor reticulatus, Rhipicephalus bursa et Ornithodoros savignyi), que ces tiques contaminées par des orbivirus lors d'un repas de sang voyaient le virus traverser leur barrière intestinale et diffuser dans l'hémolymphe vers les glandes salivaires et les ovaires. Trois semaines après nourrissage, le virus BTV8 était par exemple trouvé dans beaucoup de tissus et organes des tiques dures (contre 26 jours pour les Ornithodoros)[100] ; et le virus persistait bien dans la tique après chaque mue (« passage transstadial »). Un passage transovarien (⇒ passage du virus à la descendance) est même observé en laboratoire chez les tiques molles (alors qu'il n'a jamais pu être observé chez les culicoides vecteurs du virus Blue tongue[101]), ce qui laisse penser que certaines tiques ont un réel potentiel vectoriel[100]. Ceci expliquerait aussi l'énigme de la survie hivernale de certains virus en Europe[100]. En effet, dans les années 1970 à 2000 on admettait que le principal vecteur européen du virus BTV8 était Culicoides dewulfi, et Culicoides chiopterus[102],[103]. Or le virus ne semble pas pouvoir survivre chez ces espèces en hiver en Europe du Nord-Ouest[103]. Les capacités vectorielles d'une tique molle (Ornithodoros coriaceus) avaient d'ailleurs déjà été expérimentalement démontrées pour le BTV (Blue tongue virus) en 1985[104], avec une contamination via alimentation sur membrane artificielle ou sur des moutons ou bovins infectés. Elles s'étaient montrées capables de transmettre le virus[104].
Importance écoépidémiologique et sanitaire
[modifier | modifier le code]Alors qu'on ne connaît pas encore tous les pathogènes qu'elles peuvent inoculer (plus de 50 % des fièvres qui apparaissent en Suisse à la suite d'une morsure de tique sont encore inexpliquées[105]), les tiques semblent en augmentation dans une partie du monde, et fortement porteuses de pathogènes.
À titre d'exemple :
- chez 1 000 tiques Ixodes ricinus collectées dans une zone de loisirs boisée de Thuringe (Centre-Allemagne) des Borrelia spp., Babesia spp., Anaplasma spp., Rickettsia, Coxiella burnetii et Francisella tularensis ont été recherchées. 43,6 % des 1000 tiques étaient infectées par au moins un de ces pathogènes. Dans 8,4 % deux pathogènes différents coexistaient dans la tique (et 3 ou plus dans 1,6 % du lot étudié)[106] ;
- sur 37 zoonoses non alimentaires surveillées en priorité en France en 2006 par l’InVS, huit n'étaient transmises que par des tiques ou essentiellement par elles. I. ricinus en transmet à elle seule au moins sept, dont la maladie de Lyme, l'encéphalite à tique et la fièvre Q, qui semblent en pleine expansion et à ce titre faisant déjà partie des 11 zoonoses dont le contrôle est jugé très prioritaire en France[107] ;
- chez les oiseaux sauvages ou captifs [Psittaciformes et Passériformes, rapaces et columbiformes y semblent les plus sensibles[108] et jamais en hiver (aucun cas rapportés en novembre, décembre et janvier[108])], on parle notamment de Syndrome de mortalité des oiseaux lié aux tiques (ou ATRS pour « avian tick-related syndrome », dénomination utilisée par les anglophones)[108], qui pourrait éventuellement être une maladie émergente en Europe[108] détectée au Royaume-Uni[109] puis suspectée en France au début des années 1990[108]. Une hypothèse, à confirmer est la survenue d'une allergie (allergie croisée ?) à la salive de punaise ou plutôt de tique (Ixodes frontalis a priori selon Monks[109], spécifique des oiseaux, et largement répartie dans l'Europe, mais d'autres espèces pourraient être en cause).
Symptômes : L'état corporel semble bon, mais l'oiseau présente un abattement ou une mort brutale sans signes cliniques préalables. « Un hématome sous-cutané et un œdème sont visibles au niveau de la tête ou du cou, associés à la présence d‟une tique à proximité des lésions »(ou sur des oiseaux trouvés morts, des traces pouvant être celles de morsures de tiques sont encore visibles)[108] ; - on a récemment vu apparaître chez l'Homme une forme apparemment émergente d'allergie à la viande, d'abord en Amérique du Nord puis en Australie et en Europe. Sa cause initiale est une réaction du système immunitaire à une piqûre de tique qui inocule dans l'organisme humain une molécule présente dans la chair des mammifères[110]. On ignore si ce type d'allergie peut se déclencher chez un carnivore ou omnivore sauvage ou domestique (chat, chien).
Risques écoépidémiologiques
[modifier | modifier le code]

Des chercheurs suédois[111] ont en 2001 fait un constat identique pour les cas humains d'encéphalite à tiques en Suède : sur 30 ans, plus le climat avait été doux, plus les tiques avaient été nombreuses et plus ces encéphalites avaient également été nombreuses. Les tests sanguins faits depuis la fin des années 1950 pour chaque cas d'encéphalite diagnostiqué dans le comté de Stockholm montrent aussi très clairement que l'incidence de cette maladie a considérablement augmenté (triplement) depuis le milieu des années 1980 avec un pic en 1994 (triplement du nombre de cas humains[112]). En analysant les températures de la période 1960-1998, les chercheurs ont pu préciser que chaque augmentation de l'incidence de la maladie a été significativement liée à une combinaison de deux hivers doux avec printemps précoces ou automne doux l'année précédant le nouveau pic d'incidence. Les chercheurs ont intégré d'autres facteurs d'influence (dont l'augmentation de la population vivant en chalet d'été) dans les régions où les tiques et la maladie sont aujourd'hui endémiques, et augmentation des populations animales vectrices de tiques et/ou du microbe (facteur pour partie lié au climat).
L'accès à la vaccination contre l'encéphalite à tique en Suède (depuis 1986) et une sensibilisation accrue aux risques posés par les tiques pourraient avoir fait encore sous-estimer ces liens de cause à effet ont-ils ajouté. D'autres études, dont certaines autour de la Baltique ont montré que le réchauffement climatique ne pouvait cependant pas expliquer l'explosion du nombre de cas de maladies virales telle que l'encéphalite à tique (variante européenne)[113]
L'encéphalite à tiques est due à un Flavivirus qui comporte trois sous-types dits « européen », « sibérien » et « extrême-oriental »[114],[115]. Les cas dus au sous-type européen ont encore spectaculairement augmenté de 1995 à 2005, alors que les zones à risque ont continué à s'étendre, avec de nouveaux foyers découverts chaque année. La détection précoce de ces foyers devrait être une priorité de santé publique afin que les médecins diagnostiquent mieux et soignent plus vite leurs patients. Pour les y aider, des chercheurs ont cherché à modéliser[116] l'expansion du variant européen de cette maladie souvent mal détectée (symptômes grippaux) mais qui peut évoluer ensuite vers une méningoencéphalite aigüe et/ou une myélite qui conduit rarement à la mort, mais dont les séquelles chroniques sont invalidantes et souvent accompagnées de troubles cognitifs.
Quelques paramètres socioéconomiques sont en cause, mais ils ne peuvent expliquer l'explosion du nombre de cas humains (comme pour la maladie de Lyme). Les pullulations de tiques semblent être un facteur clé. On a montré en Amérique du Nord que ces pullulations étaient fortement associées à des changements de la structure écopaysagère des milieux forestiers. Qu'en est-il en Europe pour les tiques véhiculant les encéphalites à tiques ? Une analyse a porté (dans dix-sept provinces des Alpes au nord de l'Italie) sur d'éventuelles corrélations entre l'augmentation d'encéphalites à tique et/ou des variables climatiques et de structure de la forêt et/ou l'abondance des principaux grands vertébrés hôtes de tiques (chevreuil surtout ici), à l'aide des données disponibles pour les 40 dernières années. Aucune différence significative n'a été constatée en termes de tendance climatique entre les provinces où la maladie est apparue par rapport aux provinces où aucun cas clinique n'a été diagnostiqué (au moment de l'étude). Par contre, le meilleur modèle explicatif de l'augmentation de l'incidence de la maladie chez l'homme est celui qui intègre les changements dans la structure forestière et en particulier le ratio taillis/hauteur des forêts et les changements de densité de chevreuils.
La structure de la végétation forestière, certains changements d'affectation des sols et le nombre de chevreuils ont en effet conjointement évolué depuis 30 ans et depuis 10 ans[Quand ?], de telle sorte que les espèces-réservoirs du virus (petits mammifères) ont été fortement favorisées, entre autres par les pratiques de gestion de la faune chassable (agrainage du gibier, piégeage ou chasse des prédateurs naturels). Les auteurs pensent que ces facteurs « sont susceptibles d'être parmi les plus importants facteurs influant sur le potentiel de circulation du virus et, par conséquent, le risque d'apparition de nouveaux foyers d'encéphalites à tiques chez l'Homme en Europe occidentale. Nous pensons que notre approche sera utile pour prédire le risque TBE sur une échelle plus large », ont-ils ajouté.
Pour d'autres maladies, on a constaté que le climat et le microclimat influaient directement sur le comportement de plusieurs espèces de tiques souvent porteuses dangereuses pour l'Homme. Des chercheurs du CNRS de Marseille ont cherché à expliquer l'origine d'une petite épidémie de rickettsiose qui s'est déroulée en avril 2007 à Nîmes et pourquoi les rickettsioses avaient été en France plus nombreuses et plus graves les étés très chauds de 2003 et 2005, alors que la tique du chien (Rhipicephalus sanguineus) est surtout présente au printemps. ayant été le plus chaud dans la région depuis 50 ans, une hypothèse était que le comportement de la tique avait pu être modifié par la chaleur exceptionnelle qui a accompagné ces trois événements épidémiologiques. Et effectivement, en laboratoire[117] que si la « tique du chien » préférait réellement les chiens en temps normal, pour des raisons encore mal comprises, elle cherchait beaucoup plus à mordre l'Homme en contexte plus chaud. Des tiques de chien d'élevage, non infectées, ont été séparées en deux groupes, le premier ayant été incubé 24 heures à 40 °C, et le second à 25 °C. Puis les tiques ont été mis en présence de l'homme : « 50 % de celles incubées à 40 °C ont tenté de mordre l'homme en s'y attachant, contre aucune dans l'autre groupe ».
De plus, comme de nombreux autres organismes face aux biocides, les tiques ont montré une capacité de résistance aux acaricides[118] (notamment chez les tiques qui infestent les bovins[119], forçant les éleveurs et les producteurs d'antiparasitaires à rechercher de nouvelles molécules pesticides[120], ce qui est coûteux[121]). Des stratégies alternatives sont recherchées[122],[123].
L'importance relative des différents facteurs biotiques et abiotiques dans l'émergence et la propagation de maladies transmises par les tiques à travers l'Europe fait depuis peu l'objet d'une évaluation rigoureuse[124].
Les populations de tiques sont en effet en augmentation rapide depuis la fin du XXe siècle dans de nombreuses régions du monde, semble-t-il en raison de changements environnementaux (réchauffement climatique[125] et écopaysager ; fragmentation du paysage, espèces introduites ou invasives, diffusion de parasites par déplacement croissant d'espèces et des humains, etc.).
Actions contre les tiques
[modifier | modifier le code]Prévention
[modifier | modifier le code]
La meilleure prévention primaire est d'éviter la morsure. Il existe divers produits répulsifs, mais dont l'efficacité n'est pas toujours évidente. Après une promenade ou des activités en forêt ou dans les champs, une inspection soigneuse des vêtements et du corps permet de détecter les tiques puis de les enlever avant qu'elles aient eu le temps de transmettre la maladie de Lyme. Une personne qui est allée seule en forêt devra se faire aider pour cette inspection pour les zones difficilement visibles par elle-même comme le dos.
Des données nord-américaines et plus récemment néerlandaises ont montré que la fragmentation des forêts et la régression des prédateurs carnivores des micromammifères (renard et fouines notamment) sont des facteurs de pullulation des tiques et de risque épidémiologique accru[126],[127]. Reconstituer des continuités écopaysagères et œuvrer à la restauration d'équilibres écologiques pourraient donc, à moyen et long terme, être une mesure de prévention utile.

À l'extérieur il est conseillé, lorsque l'on va en forêt ou dans les champs :
- de porter des vêtements longs et aussi couvrants que possible[128] pour empêcher les tiques de se fixer sur la peau : les simples T-shirts et shorts doivent être évités ;
- de porter des vêtements clairs pour pouvoir localiser les tiques plus facilement ;
- de porter des chaussures montantes fermées (des bottes sont le mieux) pour éviter que les tiques ne se fixent sur la peau ;
- de porter des chaussettes « anti-tiques » dont le haut recouvre le bas du pantalon ;
- d'utiliser des répulsifs (qui sont aussi insectifuges) à base de DEET (utilisé largement depuis la fin des années 1950, notamment par l'armée américaine[129], ayant l'inconvénient d'être efficace seulement pendant quelques heures[130] et nécessitant des applications répétées) ou de p-menthane-3,8-diol[131] (ce dernier contenu dans l'huile essentielle d'eucalyptus citronné diminue significativement les morsures de tiques sans pouvoir les éliminer complètement[132]).
Les vêtements peuvent être imprégnés d'insecticides (perméthrine) ou de répulsifs. Ils sont d'une bonne efficacité[133], surtout si l'imprégnation en a été faite dès l'usine par rapport à un trempage a posteriori[134].
Il faut se débarrasser des tiques trouvées sur les vêtements, certaines pouvant survivre jusqu'à une semaine et entraîner une morsure retardée[135].
Au retour, examiner attentivement toutes les parties du corps pour y dépister les tiques éventuellement fixées à la peau puis se doucher et se changer est conseillé.
À la maison, il est possible de limiter la prolifération des tiques dans la maison et à l'extérieur :
- couper l'herbe, éliminer les feuilles mortes, refuges de larves ;
- aspirer puis boucher les creux et interstices dans les planchers, les murs ;
- inspecter les animaux au retour d'une sortie, traiter les lieux d'élevage avec des produits spécifiques ;
- empêcher l'installation facile de rongeurs dans et aux abords de la maison.
En cas de morsure
[modifier | modifier le code]

Toute tique découverte fixée doit être retirée au plus vite car le risque de contamination augmente avec la durée du contact. On estime que le risque est élevé quand la tique reste plus de 24 heures fixée sur la peau[135],[136],[137],[138].
Le crochet à tiques — vendu en pharmacie, chez les vétérinaires, etc. — permet de saisir la tique au plus près de la peau à l'aide du crochet puis tourner lentement sans tirer jusqu'à ce que la tique se décroche. Si l'on ne dispose pas de crochet à tiques, on peut utiliser une pince à bouts fins et pointus (pas une pince à épiler les sourcils qui couperait la tête de la tique) en la saisissant au plus près des pièces buccales puis en tirant sans tordre[137]. (Centres pour le contrôle et la prévention des maladies ou CDC)[139]. En l'absence de pinces, on peut utiliser un fil à coudre que l'on nouera au plus près de la peau puis on tirera dans l'axe.
Le rostre d'une tique (les pièces buccales qui sont plantées dans la peau) est entièrement recouvert d'épines qui sont implantées de façon rétrograde. Lorsqu'on utilise une autre technique que le crochet à tiques, ces épines peuvent se redresser et retenir le rostre qui risque de se casser à sa base et de rester dans la peau[140], ce qui peut être à l'origine d'une infection, et causer douleur et inflammation (nodule persistant après retrait). Cependant aucune transmission d'agents pathogènes n'est à craindre car ceux-ci sont situés dans l'abdomen. Si on tourne le corps de la tique avec un crochet prévu à cet effet, les épines reviennent autour de l'axe de rotation, et le rostre est extrait de la peau.
Il est essentiel de ne pas comprimer l'abdomen de la tique à l'occasion de son retrait, pour minimiser le risque de régurgitation de salive ; ce reflux salivaire dans la peau peut être à l'origine de phénomènes allergiques et de la transmission des microorganismes dont les tiques peuvent être les vecteurs[réf. nécessaire]. Pour la même raison, il est déconseillé d'appliquer tout produit (éther, alcool, huile…). En effet, en se sentant agressée, la tique risque de régurgiter et d'envoyer ses microbes dans l'hôte qui l'héberge. Les pinces à mors opposés et autres instruments similaires exercent une pression sur le tube digestif de la tique ; en revanche, les crochets à tiques viennent au contact du corps de la tique sans y exercer de pression excessive.[réf. nécessaire]
Une fois la tique retirée, on peut utiliser un système d'aspiration pour extraire, à partir de la morsure, une goutte de lymphe ou de sang (éventuellement infectés). Cela atténue au moins les démangeaisons. Ne pas oublier de bien désinfecter.
La surveillance de la plaie est préconisée pendant trois semaines. La présence d'une auréole rouge qui évolue (érythème migrant), la survenue d'un état grippal doit faire craindre la survenue d'une maladie de Lyme demandant un traitement spécifique. Dans les régions à haute endémie, la mise systématique sous antibiotiques (doxycycline) peut être préconisée[141].
D'autres méthodes de retrait ont été proposées dans le but d'« étouffer » la tique et d'en provoquer la chute spontanée (application de vernis à ongles, d'essence) ; elles sont inefficaces[140].
Pour les animaux
[modifier | modifier le code]Les conséquences des infestations sur les animaux :
Les infestations à tique sont importantes et de plus en plus fréquentes pour les animaux sauvages et domestiques, elles peuvent transmettre plusieurs maladies.
Les tiques semblent être les uniques vecteurs d'une maladie parasitaire du sang appelée piroplasmose (ou Babésiose) due à un protozoaire (Babesia sp., ex : B. canis, B. microtii). La piroplasmose atteint les canidés, les équidés, les bovidés… Le protozoaire parasite les globules rouges dans lesquels il se multiplie et les détruit lorsqu'il s'en libère. La piroplasmose entraîne une anémie avec un ictère pâle et généralement une forte fièvre. Dans les stades avancés de la maladie, une coloration brun foncé des urines peut être notée, due à la bilirubine issue de la dégradation de l'hémoglobine libérée dans le sang par la destruction des globules rouges. La piroplasmose est une maladie potentiellement mortelle.
Trois moyens de luttes contre les parasites :
- action pour rompre le cycle externe ⇒ épandage de produits acaricides. Ils semblent peu efficaces en raison du réservoir naturel ;
- action sur l’animal pour détruire le parasite ⇒ douchage, balnéation avec une solution acaricide ;
- défragmenter les écosystèmes et y laisser les prédateurs vivre, pour y restaurer des équilibres écologiques, de manière à limiter les pullulations de tiques.
Utilisations à des fins médicales
[modifier | modifier le code]Plusieurs équipes scientifiques à travers le monde ont identifié, dans la salive de diverses espèces de tiques, des protéines capables d’influencer la coagulation (composés hémostatiques, notamment chez les tiques dures qui peuvent se nourrir jusqu'à deux semaines[142]), l’inflammation et la réponse immunitaire. Les gènes correspondants ont été clonés à partir de l'ADN des tiques et des protéines recombinantes ont été produites. Les scientifiques examinent la possibilité de les utiliser comme agents thérapeutiques dans le traitement des maladies qui font intervenir une composante inflammatoire ou auto-immunitaire. Pour certaines d’entre elles, les essais sur animaux sont encourageants[143].
Recherche
[modifier | modifier le code]
La recherche, à l'origine essentiellement axée sur les moyens de lutte pour détruire les tiques sur les animaux domestiques, s'est ensuite intéressée aux germes pathogènes véhiculés par ces ectoparasites, au fur et à mesure de leur découverte. Pendant très longtemps, la recherche s'est cantonnée au seul domaine vétérinaire, pour s'ouvrir au domaine de la médecine humaine à partir surtout de la deuxième moitié du XXe siècle. D'abord, avec les très nombreux travaux européens sur les virus à l'origine de l'encéphalite à tiques (RSSE et CEE), ainsi que les études américaines sur les rickettsies à l'origine de la fièvre pourprée des Montagnes Rocheuses, puis avec ceux sur les borrélies responsables de la maladie de Lyme, aux États-Unis à partir du milieu des années 1970, en Europe ensuite. Elle tente maintenant de clarifier les systèmes complexes multi-vecteurs, multi-hôtes, avec la diversité des modes de transmission dans lesquelles les tiques interagissent et évoluent, systèmes qui se modifient du fait de la mondialisation des échanges et des modifications environnementales induites par l'homme (dont climatiques et en termes d'écologie du paysage). Le risque d'émergence accrue de maladies et pathogènes transmis par les tiques est maintenant avéré, certaines tiques faisant presque figure de candidat idéal pour leur vectorisation. Des progrès importants ont été permis par l'établissement d'une méthode efficace d'extraction de l'ADN de tiques[144] et par les techniques de détection globale de l'ADN des agents bactériens abrités dans les tiques (ex : DEA de Lénaig Halos, 2002 / PCR-TTGE ou Temporal Température Gel Electrophoresis). L'ADN bactérien est maintenant détecté amplification du gène de l'ARN16S (commun à tous les procaryotes) grâce à des amorces universelles. Ceci permet notamment de mieux comprendre et évaluer les risques de co-infection par plusieurs micropathogènes[145].
On cherche notamment à comprendre et à modéliser ce qui peut expliquer les rapides pullulations de tiques, localement observées dans le monde, les motifs de variations spatio-temporelles, le portage de bactéries ou autres microbes par ces tiques, ainsi que leur rôle éco-épidémiologique. Ceci change l'incidence des maladies transmises par les tiques dans cette région (données non analysées à ce jour[Quand ?]). Ces travaux sont nécessaires pour modéliser les risques d'émergence de pathogènes et maladies à tiques chez les animaux domestiques, sauvages et chez l'Homme, et pour proposer des solutions alternatives et efficaces aux pesticides pour leur contrôle (certaines tiques, se sont montrées capables de très rapidement développer des résistances par adaptation sélective à tous les types d'acaricides utilisés contre elles).
Un espoir est celui de comprendre quels sont les parasites et régulateurs naturels des tiques et pourquoi ils ont régressé ou ne sont plus efficaces dans le contrôle de celles-ci. Une espèce au moins a été trouvée presque partout dans le monde, et est étudiée pour ces raisons Ixodiphagus hookeri.
En France
[modifier | modifier le code]L'enquête de Cuisance et Rioux (2004) a conclu qu'il n'existait en France en 2003 plus que deux spécialistes actifs en systématique des tiques (partis à la retraite depuis). Cependant, pour les tiques de la faune de France, transposable aux pays voisins de même type climatique, l'ouvrage de Claudine Pérez-Eid paru en 2007 est très complet[2], proposant des clés d'identification et des schémas de l'ensemble des espèces présentes en France.
Quelques museums et laboratoires disposent de collections de tiques, telle que celle de Montpellier (300 espèces et 7 634 échantillons)[146].
Le groupe Tiques et maladies à tiques[147] a été créé en septembre 2004 au sein du REID (Réseau écologie des interactions durables), créé par l'INRA en 1993 pour mieux étudier, collaborativement, l'écologie, l'évolution adaptative et la génétique[148] des tiques et des pathogènes qu'elles véhiculent ainsi que les interactions tique-environnement » et les problèmes d'échantillonnage, évaluation des populations et de leurs caractéristiques génétiques, d'élevage de laboratoire et d'analyse d'échantillons. Ce groupe regroupait, mi-2008, 46 membres de dix grands établissements scientifiques (INRA, CNRS, CIRAD, IRD, MNHN, Tour du Valat, École nationale vétérinaire de Maison-Alfort, université de Bourgogne, université de Rennes, université de Strasbourg) ainsi qu'un médecin généraliste et des vétérinaires, dont l'un représentant la Société nationale des groupements techniques vétérinaires (SNGTV). Il a notamment (avec l'INSERM et au sein de l'unité mixte de recherche BIPAR, avec l'Institut Pasteur, avec l'aide financière de l'Union européenne[149] commencé à explorer le microbiote intestinal (bactéries, virus et protozoaires) de tiques provenant d'Alsace et des Ardennes françaises, grâce au séquençage haut-débit.
En 2018, ce travail avait déjà permis de découvrir dans l'intestin de ces tiques de nombreux organismes inconnus ou inattendus (« de nombreux nouveaux virus identifiés dans ces tiques sont en cours de caractérisation » précisait l'INRA en 2018) tels que[150],[151],[152],[153],[154],[155] :
- la bactérie Borrelia miyamotoi, connue pour être associée à des fièvres sévères ;
- la bactérie Neoehrlichia mikurensis également associée à des fièvres sévères ;
- plusieurs bactéries du genre Bartonella, jusqu'alors non découvertes, associées à une tique en France, et ici également retrouvées dans le sang de 6 personnes dans un groupe de 66 personnes piqués par des tiques (personnes choisies de manière à couvrir toutes les régions de France) ;
- trois souches de Bartonella henselae (agent responsables chez l'Homme de la maladie des griffes du chat, également retrouvées dans le tube digestif des tiques étudiées) ;
- plusieurs parasites nouveaux du genre Babesia et Theileria (peut être pathogènes pour l'Homme) ;
- le virus Eyach suspecté d'être zoonotique et dangereux pour l'Homme ;
- la bactérie Bartonella doshiae, qui était connue chez les campagnols et mulots, mais ici aussi retrouvée dans le sang de patients mordus par des tiques ;
- la bactérie Bartonella tribocorum, qui était connue chez les rats, mais ici aussi retrouvée dans le sang de patients mordus par des tiques ;
- la bactérie Bartonella schoenbuchensis, qui était connue chez les cervidés, mais ici aussi retrouvée dans le sang de patients mordus par des tiques.
En 2016-2017, ces travaux ont aussi pu confirmer la fréquence des infections chez l'Homme, qui est même « la règle, plutôt que l'exception »[156],[157]. Un autre projet, OSCAR (2012-2016), met au point un outil de cartographie du risque infectieux, par simulation informatique (à l'échelle du paysage agricole)[158]. Un projet XENOBIO-TICK vise à produire de nouveaux acaricides, alors que le projet Vactix vise un vaccin anti-tique qui doit être testé sur des ovins[158].
Programmes français de sciences participatives
[modifier | modifier le code]CiTIQUE [159] est un programme de recherche participative partenarial qui vise à mieux comprendre l'écologie des tiques et les maladies qu'elles transmettent, dont la maladie de Lyme. CiTique fait travailler ensemble, et à toutes les étapes du projet des citoyens et des chercheurs. Ce projet est porté par des équipes de l'INRA[160] missionnées sur l'étude des tiques et des maladies transmises par elles, depuis le début des années 2000, CPIE[161] et ANSES[162]. Le projet est soutenu par le ministère de la santé[163] dans le cadre du plan Lyme et un travail initié en 2013 d'étude des pathogènes véhiculés par la tique au sein de son écosystème microbien, avec les interactions qu'il y développe (Pathobiome)[158]. L'INRA inscrit son travail dans une approche One Health- EcoHealth[158].
Dès [164] une application pour smartphone et un site internet (Signalement tique[165]) ont vu le jour, portés par l'INRA, aujourd'hui devenue Institut national de recherche pour l'agriculture, l'alimentation et l'environnement (INRAE). Un engouement important a eu lieu puisqu'en moins de deux mois (entre le et le ), l'application a été téléchargée 20 520 fois et a permis le signalement de 3 100 piqûres (animaux et humains compris)[166]. Une fois la piqûre de tique signalée, le citoyen peut d'envoyer la tique piqueuse à un laboratoire (adresse unique pour toute la France : Laboratoire Tous chercheurs, Inra Nancy Champenoux – rue d'amance 54280 Champenoux).
Cette collecte sans précédent en France, d'individus piqueurs, sur humains et sur animaux doit fournir une collection unique de données et d'échantillons. Cette « tiquothèque » sera ouverte à d'autres chercheurs travaillant sur les tiques, ils gagneront ainsi du temps et de l'efficacité. Déjà, d'autres projets de recherche s'appuient sur les données de CiTIQUE[167], comme Ohticks[168] (coordonné par l'Inra et visant à mieux connaitre les agents pathogènes transmis par les tiques à l'Homme et aux animaux, ainsi que les risques de co-infection pour proposer des tests de diagnostic plus efficaces ; en 2018 selon l'INRA « La moitié des microorganismes vectorisés par les tiques ne sont pas connus et ils sont peut-être impliqués dans les maladies transmises » mais le séquençage haut débit peut aider à identifier quels sont ces nouveaux microbes), TéléTiQ[169], ou [GenIric] (visant à séquencer exhaustivement les génomes d'Ixodes ricinus).
D'autres actions sont prévues dans ce cadre dont l'ouverture d'un laboratoire spécialement aménagé, appelé « Tous chercheurs », où les citoyens pourront venir travailler aux côtés des scientifiques afin de parfaire leurs connaissances du sujet et de débattre des questions auxquelles tentent de répondre par la science les chercheurs. Une communication vers les citoyens doit améliorer les réflexes de prévention et la culture partagée au sujet des tiques.
Historique
[modifier | modifier le code]La connaissance des tiques par les auteurs anciens semble avoir été très lacunaire. Longtemps masculin, le nom tique est devenu féminin dans le langage courant.
Curieusement, alors que les poux et puces sont bien décrits par les chroniqueurs et médecins de l'antiquité à nos jours, et que de nombreuses potions et recettes existaient pour s'en débarrasser, les tiques semblent très rarement évoquées par les textes du passé.
Plusieurs dictionnaires anciens évoquent la tique aussi appelée « ricin » comme un insecte infectant les chiens et les bœufs.
- « Tique, ou Tiquet, Ricinus, Croton vermis canes et boues infestans, non habens exitum excrementi. »[170] ;
- « Insecte noirâtre qui s'attache aux oreilles des chiens, des bœufs, &c. La tique crève après s'être gorgée de sang. » pour l'Académie française en 1762[171],
Émile Littré en précise dans le dernier quart du XIXe siècle la définition comme suit, en signalant que la tique s'attaque aussi au mouton et à d'autres animaux, mais sans évoquer l'homme (alors que la définition du pou dans le même dictionnaire le cite très explicitement).
- « Un des noms vulgaires donnés aux espèces du genre ixode, et surtout à l'ixode ricin qui s'attache aux chiens, aux bœufs, aux moutons et autres animaux (les ixodes ne sont pas des insectes, mais des arachnides).
- BUFF., Ois. t. XII, p. 92: « Ils [certains oiseaux] se posent sur les bœufs et sur les vaches pour manger les tiques, les vers et les insectes nichés dans le poil de ces animaux »
- « Soûl comme une tique, très soûl, vu que la tique se gorge de sang et semble immobile ensuite. »[172]
La tique du chien était aussi autrefois aussi appelée « Rézée » ou « lagast » dans le Languedoc[173]. Le Nouveau dictionnaire d'histoire naturelle, appliquée aux arts, à l'agriculture, à l'économie rurale et domestique, à la médecine, etc., en 1819 (p. 295 du tome XXIX) dit en parlant de la graine de ricin dont on extrait l'huile bien connue : « Nos romains l'appellent ricinus parce qu'elle ressemble au ricinus, espèce de pou qui tourmente les moutons » (Le ricin était aussi appelé « Croton », nom également donné aux tiques à cette époque.)
Les auteurs anciens ne font curieusement pas allusion au fait que des tiques mordaient les hommes. Ils les citent comme affectant le bétail, les oiseaux et les chiens.
Dans la culture
[modifier | modifier le code]Dans Le Parfum, histoire d'un meurtrier, Patrick Suskind compare de manière particulièrement suggestive à une tique le personnage de Jean-Baptiste Grenouille qui choisit de vivre, non guidé par la raison mais « de façon végétative » :
« […] comme la tique sur son arbre, à laquelle pourtant la vie n'a rien d'autre à offrir qu'une perpétuelle hibernation. La petite tique toute laide, qui donne à son corps couleur de plomb la forme d'une boule, afin d'exposer le moins de surface possible au monde extérieur ; qui rend sa peau dure et sans faille, pour ne rien laisser filtrer, pour qu'il ne transpire absolument rien d'elle au-dehors. La tique, qui se fait délibérément petite et terne, pour que personne ne la voie et ne l'écrase. La tique solitaire, concentrée et cachée dans son arbre, aveugle, sourde et muette, tout occupée, pendant des années, à flairer sur des lieux à la ronde le sang des animaux qui passent et qu'elle n'atteindra jamais par ses propres moyens. La tique pourrait se laisser tomber. Elle pourrait se laisser choir sur le sol de la forêt et, sur ses huit minuscules pattes, se traîner de quelques millimètres dans un sens ou dans l'autre pour se disposer à mourir sous une feuille, ce ne serait pas une perte, Dieu sait ! Mais la tique, butée, bornée, et répugnante, reste embusquée, et vit, et attend. Attend jusqu'à ce qu'un hasard extrêmement improbable lui amène le sang juste sous son arbre, sous la forme d'un animal. Et c'est alors seulement qu'elle sort de sa réserve, se laisse tomber, se cramponne, mord et s'enfonce dans cette chair inconnue[174]… »
Dans la téléfilm L'Abécédaire de Gilles Deleuze, à la lettre "A - comme Animal", Gilles Deleuze évoque la tique. Il reprend ainsi, sans en faire mention, l'exemple qu'utilise Jacob von Uexküll, dans son livre Milieu animal et milieu humain, pour expliquer la notion d'Umwelt :
« […] la question c'est "quel rapport avez-vous avec l'animal ?" […] C'est vrai que je suis fasciné par des bêtes comme les araignées, les tiques, les poux, tout ça… c'est aussi important que les chiens et les chats. Et c'est des rapports avec les animaux aussi, quelqu'un qui a des tiques, des poux… qu'est-ce que ça veut dire tout ça… c'est des rapports avec les animaux très actifs. […] Qu'est-ce qui me frappe chez un animal ? La première chose qui me frappe, je crois, c'est le fait que tout animal a un monde […]. Il y a des tas d'humains qui n'ont pas de monde, ils vivent la vie de tout le monde […]. Les animaux, ils ont des mondes, un monde animal c'est quoi ? C'est parfois extrêmement restreint, et c'est ça qui m'émeut ! […] La tique répond ou réagit à trois choses, trois excitants, un point c'est tout. […]. Elle tend vers l’extrémité d'une branche d'arbre, attirée vers la lumière […]. Elle sent, la tique sent la bête qui passe sous sa branche, ça fait deuxième excitant, lumière et puis, odeur […]. Et puis […] elle va chercher la région la moins fournie en poils, donc là un excitant tactile, et elle s'enfonce sous la peau. Le reste, elle s'en fout complètement, dans une nature fourmillante, elle extrait, elle sélectionne trois choses […], c'est ça qui fait un monde[175]. »
Dans l'entre-deux-guerres mondiales, les fascistes italiens appelaient les membres de l'extrême-gauche, communistes et anarchistes, les tiques. Ce surnom a été repris lors d'une opération de la police italienne visant les anarchistes, l'opération Ixodida[176].
Notes et références
[modifier | modifier le code]Notes
[modifier | modifier le code]- Dans la plupart des pays, notamment anglo-saxons, la vitamine connue en France comme « vitamine B8 », la biotine, est dénommée « vitamine B7 »[70].
Références
[modifier | modifier le code]- Guglielmone, Robbins, Apanaskevich, Petney, Estrada-Pena, Horak, Shao & Barker, 2010 : The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: a list of valid species names Zootaxa, n. 2528, p. 1–28.
- Claudine Pérez-Eid, « Les tiques : identification, biologie, importance médicale et vétérinaire » (Coll. Monographies de microbiologie), Lavoisier, 2007.
- Alejandro Cabezas-Cruz et James J. Valdés, « Are ticks venomous animals? », sur Frontiers in Zoology, (ISSN 1742-9994, PMID 25006341, PMCID PMC4085379, DOI 10.1186/1742-9994-11-47, consulté le ), p. 47
- « Audition plénière - Borréliose de Lyme - Audition commune de cadrage épidémiologique et biologique de la borréliose de Lyme - Alain MILON », sur videos.senat.fr (consulté le ).
- Sonenshine, D. E. 1991. Biology of ticks. Vol. 1. Oxford University Press, New York.
- « Moustique tigre, tique... Les (sales) petites bêtes qui montent », sur Nouvel Obs, .
- (en) El Shoura SM (1988) Fine structure of the sight organs in the tickHyalomma (Hyalomma) dromedarii (Ixodoidea: Ixodidae). Experimental & applied acarology, 4(2), 109-116. (résumé).
- Kaltenrieder, M., Labhart, T., & Hess, E (1989) Spectral sensitivity, absolute threshold, and visual field of two tick species, Hyalomma dromedarii andAmblyomma variegatum. Journal of Comparative Physiology A, 165(2), 155-164 (résumé).
- Leonovich SA (1985) [Orientational behavior of the ixodid tick Hyalomma asiaticum under desert conditions]. Parazitologiia, 20(6), 431-440 (résumé).
- JC George, Les Tiques - Ixodidae, voir le chap. "Écologie et éthologie". Consulté 2014-04-24.
- Diaporama du groupe Tiques de Flickr, montrant des dizaines d'espèces et stades différents.
- (de) Hopf H. « Neurologische Kompilationen nach Zeckenkontakt » Dtsch. Artzebl. 1980, 77, p. 2063-2066.
- Sonenshine, D. E., Silverstein, R. M., Plummer, E., West, J. R., & McCullough, T. (1976). 2,6-Dichlorophenol, the sex pheromone of the Rocky Mountain wood tick, Dermacentor andersoni Stiles and the American dog tick, Dermacentor variabilis (Say). Journal of Chemical Ecology, 2(2), p. 201-209.
- Borges, L. M., Eiras, Á. E., Ferri, P. H., & Lôbo, A. C. C. (2002). The role of 2, 6-dichlorophenol as sex pheromone of the tropical horse tick Anocentor nitens (Acari: Ixodidae), Experimental and Applied Acarology, 27(3), p. 223-230.
- [PDF] De Bruyne, M., & Guerin, P. M. (1994). Isolation of 2, 6-dichlorophenol from the cattle tick Boophilus microplus : receptor cell responses but no evidence for a behavioural response, journal of insect physiology, 40(2), p. 143-154.
- Site labellisé HON sur les maladies a tiques http://www.maladies-a-tiques.com.
- Site sur les tiques du groupe Baxter http://www.latique.ch.
- Page sur les tiques en collaboration avec le Conservatoire Botanique National des Pyrénées http://mycologie.catalogne.free.fr/Tiques.htm.
- source : Gwenaël Vourc'h, Unité d'épidémiologie animale, INRA, St Genès Champanelle, France.
- (en) Enrique Peñalver, Antonio Arillo, Xavier Delclòs, David Peris, David A. Grimaldi, Scott R. Anderson, Paul C. Nascimbene et Ricardo Pérez-de la Fuente, « Parasitised feathered dinosaurs as revealed by Cretaceous amber assemblages », Nature Communications, vol. 8, no 1, (DOI 10.1038/s41467-017-01550-z).
- Zhang X.Z., Meltzer M.I., Pena C.A., Hopkins A.B., Wroth L. & Fix A.D. (2006) Economic impact of Lyme disease. Emerging Infectious Diseases, 12, 653-660.
- Kurtenbach, K et al. 2006. Fundamental processes in the evolutionary ecology of Lyme borreliosis. Nature Rev. Microbiol. 4: 660-669.
- J.L. Corn, N. Barré, B. Thiébot, T.E Creekmore, G.I. Garris et V.F. Nettle, « A study of the potential role of cattle egrets Bubulcus ibis (Ciconiformes : Ardeidae) in the dissemination of Amblyomma variegatum (Acari: Ixodidae) in the Eastern Carribean », J. Med. Entomol., vol. 30, , p. 1029-1037.
- La vie des tiques Voir paragraphe reproduction.
- Arthur, D. R. 1961. Ticks and diseases. Row, Peterson and Company, New York.
- Francischetti, I. M., Sa-Nunes, A., Mans, B. J., Santos, I. M., and Ribeiro, J. M. (2009) The role of saliva in tick feeding. Front. Biosci. 14, 2051–2088. doi: 10.2741/3363
- Mans B.J (2011) Evolution of vertebrate hemostatic and inflammatory control mechanisms in blood-feeding arthropods. J. Innate Immun. 3, 41–51. doi: 10.1159/000321599
- Kazimirova, M., and Stibraniova, I. (2013) Tick salivary compounds: their role in modulation of host defences and pathogen transmission. Front. Cell. Infect. Microbiol. 3:43. doi: 10.3389/fcimb.2013.00043
- Wikel S (2013) Ticks and tick-borne pathogens at the cutaneous interface: host defenses, tick countermeasures, and a suitable environment for pathogen establishment. Front. Microbiol. 4:337. doi:10.3389/fmicb.2013.00337
- Valdes, J. J. (2014) Antihistamine response: a dynamically refined function at the host-tick interface. Parasit. Vectors 7:491. doi: 10.1186/s13071-014-0491-9
- Kotal, J., Langhansova, H., Lieskovska, J., Andersen, J. F., Francischetti, I. M., Chavakis, T., et al. (2015) Modulation of host immunity by tick saliva. J. Proteomics 128, 58–68. doi: 10.1016/j.jprot.2015.07.005
- Chmelar, J., Kotal, J., Kopecky, J., Pedra, J. H. & Kotsyfakis, M. (2016) All for one and one for all on the tick-host battlefield. Trends Parasitol. 32, 368–377. doi: 10.1016/j.pt.2016.01.004
- Šimo, L., Kazimirova, M., Richardson, J., & Bonnet, S. I. (2017). The essential role of tick salivary glands and saliva in tick feeding and pathogen transmission. Frontiers in cellular and infection microbiology, 7, 281.
- Brossard M & Wikel S.K (1997) Immunology of interactions between ticks and hosts. Medical and Veterinary Entomology 11: 270-276.
- Marchal C (2009) Inflammation cutanée et borréliose de Lyme: rôle de l'immunité innée et de la tique dans la transmission à l'homme de Borrelia burgdorferi | Thèse de Doctorat, Strasbourg (résumé).
- Štibrániová, I., Lahová, M., and Bartíková, P. (2013). Immunomodulators in tick saliva and their benefits. Acta Virol. 57, 200–216. doi: 10.4149/av_2013_02_200
- Ribeiro J.M.C (1982) The antiserotonin and antihistamine activities of salivary secretion of Rhodnius prolixus. J. Insect Physiol., 28: 69–75.
- Ribeiro J.M.C. & Spielman A (1986) Ixodes dammini: Salivary anaphylatoxin inactivating activity. Exp. Parasitol., 62: 292–297 (résumé)
- Grillon A (2017) Borréliose de Lyme: rôle de l’interface cutanée et du microbiome dans la physiopathologie de la maladie |Thèse de Doctorat, Strasbourg (résumé).
- Lenka Grunclov, Hélène Fouquier, Václav Hyps et Petr Kopásek, Lysozyme from the gut of the soft tick Ornithodoros moubata: the sequence, phylogeny and post-feeding regulation ; Developmental & Comparative Immunology Volume 27, Issue 8, September 2003, Pages 651-660 doi:10.1016/S0145-305X(03)00052-1 (Résumé).
- Susanne Neuperta, Reinhard Predel, William K. Russellc Ronald Daviesd Patricia V. Pietrantonioe et Ronald J. Nachman, Identification of tick periviscerokinin, the first neurohormone of Ixodidae: Single cell analysis by means of MALDI-TOF/TOF mass spectrometry ; Biochemical and Biophysical Research Communications Volume 338, Issue 4, 30 December 2005, Pages 1860-1864 doi:10.1016/j.bbrc.2005.10.165.
- S. Latour, Biologie des tiques et rôle pathogène. Cahier du Vétomecum, 1997, 3-8.
- Aeschlimann, A. (1981). The role of hosts and environment in the natural dissemination of ticks. Studies on a Swiss population of Ixodes ricinus L., 1758. Review of advances in parasitology, 859-869.
- Sonenshine, D.E. (1993). Biology of ticks, Oxford: Oxford University Press.
- Degeilh, B. (2007). Données fondamentales à la base des mesures préventives. Médecine et maladies infectieuses, doi: 10.1016/j.medmal.2006.01.031.
- France Lyme, consulté 2015-07-11.
- Claire Asher ; “Lyme disease–carrying ticks are now in half of all U.S. counties” mis en ligne le 18 janvier 2016; DOI: 10.1126/science.aae0246.
- « Edition du soir Ouest France », sur www.ouest-france.fr (consulté le )
- Bahrani, A., White, A., Bitzer, L., & Suarez, A. (2017). Capture-Mark-Recapture of Ticks in Southeastern Virginia. |résumé/posters symposium 2017).
- Les poulets mangent les tiques sur le bétail, Radios Rurales Internationales, Avril 1999.
- Corn, J. L., Barré, N., Thiebot, B., Creekmore, T. E., Garris, G. I., & Nettles, V. F. (1993). Potential role of cattle egrets, Bubulcus ibis (Ciconiformes: Ardeidae), in the dissemination of Amblyomma variegatum (Acari: Ixodidae) in the eastern Caribbean. Journal of medical entomology, 30(6), 1029-1037. (résumé).
- Voir (PDF d'un PowerPoint de 18 diapositives, fait par le Dr Kathrin Hartelt (RPS, Landesgesundheitsamt), pour la DRASS d'Alsace).
- (en) Ostfeld RS, Price A, Hornbostel VL, Benjamin MA, Keesing F, « Controlling Ticks and Tick-borne Zoonoses with Biological and Chemical Agents » BioScience 2006;56(5):383-394.
- (en) Keesing K, Brunner J, Duerr S, Killilea M, LoGiudice K, Schmidt K, Vuong H & Ostfeld RS, « Hosts as ecological traps for the vector of Lyme disease » Proc. R. Soc. B. 2009;276:3911-3919.
- Dinsmore J.J (1973) Foraging success of cattle egrets, Bubulcus ibis. American Midland Naturalist, 242-246.
- F Gigon F, Biologie d'Ixodes ricinus L sur le Plateau Suisse - Une contribution à l'écologie de ce vecteur Faculté des Sciences de Neuchâtel, thèse de doctorat ès sciences, 1985 [PDF].
- (en) Davis AJ (1986) « Bibliography of Ixodiphagini (Hymenoptera, Chalcidoidea, Encyrtidae), parasites of ticks (Acari, Ixodidae), with notes on their biology » Tijdschrift voor Entomologie 1986;129(6):181-190.
- (en) Mwangi EN, Hassan SM, Kaaya GP, Essuman S, « The impact of Ixodiphagus hookeri, a tick parasitoid, on Amblyomma variegatum (Acari: Ixodidae) in a field trial in Kenya » Exp and Appl Acar. 1997;21(2):117-126. (résumé).
- (en) Howard AFV, N'Guessan R, Koenraadt CJM, Asidi A, Farenhorst M, Akogbéto M, Thomas MB, Knols BGJ. & Takken W, « The entomopathogenic fungus Beauveria bassiana reduces instantaneous blood feeding in wild multi-insecticide-resistant Culex quinque-fasciatus mosquitoes in Benin, West Africa » Parasites & Vectors 2010;3:87.
- (en) Thaler JO, Duvic B, Givaudan A & Boemare N, « Isolation and entomotoxic properties of the Xenorhabdus nematophilus F1 lecithinase » Appl Environ Microbiol. 1998;64(7):2367-73.
- (en) Nguyen KB, A new nematode parasite of mole crickets. Dissertation to the graduate school, Université de Floride, 1988.
- (en) Veronika Burešová, Zdeněk Franta et Petr Kopáček, « A comparison of Chryseobacterium indologenes pathogenicity to the soft tick Ornithodoros moubata and hard tick Ixodes ricinus » Journal of Invertebrate Pathology 2006;93(2):96-104. DOI 10.1016/j.jip.2006.05.006 (Résumé).
- Exemple : Pages des symposiums internationaux de Potsdam, qui se réunissent depuis 1990 à Potsdam sur les maladies à tiques, avec un objectif de veille et de formation continue pour les médecins allemands (voir actes de 2001, 2003, 2005, 2007). Voir aussi les travaux du groupe tique et du CNR Borréliose en France, et de nombreuses études conduites autour de la Baltique ou aux États-Unis.
- Brownstein, J.S., Holford, T. et Fish, D. (2003). A climate-based model predicts the spatial distribution of the Lyme disease vector Ixodes scapularis in the United States. Environmental Health Perspectives 111(9), 1152-1157.
- Ostfeld R.S, Canham C.D, Oggenfuss K, Winchcombe R.J & Keesing F (2006). Climate, deer, rodents, and acorns as determinants of variation in Lyme-disease risk. PLoS biology, 4(6), e145 |URL http://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.0040145.
- Ostfeld R.S (2002) Ecological webs involving acorns and mice. Oak forest ecosystems: ecology and management for wildlife. Johns Hopkins University Press, Baltimore, MD, 196-214.
- Ostfeld R.S, Schauber E.M, Canham C.D, Keesing F, Jones C.G & Wolff J.O (2001). Effects of acorn production and mouse abundance on abundance and Borrelia burgdorferi infection prevalence of nymphal Ixodes scapularis ticks. Vector Borne and Zoonotic Diseases, 1(1), 55-63 |URL : https://www.academia.edu/download/42501147/Effects_of_Acorn_Production_and_Mouse_Ab20160209-24834-crq8d.pdf.
- Schauber, E. M., Ostfeld, R. S., & Evans Jr, A. S. (2005). What is the best predictor of annual Lyme disease incidence: weather, mice, or acorns?. Ecological Applications, 15(2), 575-586 | URL:http://izt.ciens.ucv.ve/ecologia/Archivos/ECO_POB_2005/ECOPO6_2005/Schauber%20et%20al%202005.pdf.
- Olivier Duron et Yuval Gottlieb, « Convergence of Nutritional Symbioses in Obligate Blood Feeders », Trends in Parasitology, vol. 36, no 10, , p. 816–825 (ISSN 1471-4922, DOI 10.1016/j.pt.2020.07.007, lire en ligne, consulté le )
- (en) « Biotin », sur MedlinePlus – National Institute of Health (consulté le ).
- « Chez les tiques, vivre de sang grâce à des bactéries », sur CNRS, (consulté le ).
- (en) Olivier Duron, Olivier Morel, Valérie Noël, Marie Buysse, Florian Binetruy et al., « Tick-bacteria mutualism depends on B vitamin synthesis pathways », Current Biology, vol. 28, no 12, , p. 1896–1902.e5 (DOI 10.1016/j.cub.2018.04.038).
- (en) Olivier Duron, Florian Binetruy, Valérie Noël et Julie Cremaschi, « Evolutionary changes in symbiont community structure in ticks », Molecular Ecology, vol. 26, no 11, , p. 2905–2921 (ISSN 1365-294X, DOI 10.1111/mec.14094, lire en ligne, consulté le )
- (en) Florian Binetruy, Marie Buysse, Quentin Lejarre et Roxanne Barosi, « Microbial community structure reveals instability of nutritional symbiosis during the evolutionary radiation of Amblyomma ticks », Molecular Ecology, vol. 29, no 5, , p. 1016–1029 (ISSN 1365-294X, DOI 10.1111/mec.15373, lire en ligne, consulté le )
- Itai Lalzar, Yael Friedmann et Yuval Gottlieb, « Tissue tropism and vertical transmission of Coxiella in Rhipicephalus sanguineus and Rhipicephalus turanicus ticks », Environmental Microbiology, vol. 16, no 12, , p. 3657–3668 (ISSN 1462-2920, PMID 24650112, DOI 10.1111/1462-2920.12455, lire en ligne, consulté le )
- (en) Marie Buysse, Olivier Plantard, Karen D. McCoy et Olivier Duron, « Tissue localization of Coxiella-like endosymbionts in three European tick species through fluorescence in situ hybridization », Ticks and Tick-borne Diseases, vol. 10, no 4, , p. 798–804 (ISSN 1877-959X, DOI 10.1016/j.ttbdis.2019.03.014, lire en ligne, consulté le )
- (en) Melina Garcia Guizzo, Luís Fernando Parizi, Rodrigo Dutra Nunes et Renata Schama, « A Coxiella mutualist symbiont is essential to the development of Rhipicephalus microplus », Scientific Reports, vol. 7, no 1, , p. 17554 (ISSN 2045-2322, PMID 29242567, PMCID PMC5730597, DOI 10.1038/s41598-017-17309-x, lire en ligne, consulté le )
- Michael Ben-Yosef, Asael Rot, Mustafa Mahagna et Einat Kapri, « Coxiella-Like Endosymbiont of Rhipicephalus sanguineus Is Required for Physiological Processes During Ontogeny », Frontiers in Microbiology, vol. 11, , p. 493 (ISSN 1664-302X, PMID 32390951, PMCID 7188774, DOI 10.3389/fmicb.2020.00493, lire en ligne, consulté le )
- Olivier Duron, Karim Sidi-Boumedine, Elodie Rousset et Sara Moutailler, « The Importance of Ticks in Q Fever Transmission: What Has (and Has Not) Been Demonstrated? », Trends in Parasitology, vol. 31, no 11, , p. 536–552 (ISSN 1471-4922, DOI 10.1016/j.pt.2015.06.014, lire en ligne, consulté le )
- (en) Olivier Duron, « The IS1111 insertion sequence used for detection of Coxiella burnetii is widespread in Coxiella-like endosymbionts of ticks », FEMS Microbiology Letters, vol. 362, no 17, (ISSN 0378-1097, DOI 10.1093/femsle/fnv132, lire en ligne, consulté le )
- (en) Feuente, J. de la (2003) The fossil record and the origin of ticks (Acari: Parasitiformes: Ixodida) in "Experimental and Applied Acarology" 29, 331-334. Résumé ; Consulté 2010/03/30.
- (en) B.J. Mans, D. de Klerk, R. Pienaar et A.A. Latif, « Nuttalliella namaqua: a living fossil and closest relative to the ancestral tick lineage: implications for the evolution of blood-feeding in ticks », PLoS One, vol. 6, no 8, , e23675 (DOI 10.1371/journal.pone.0023675, lire en ligne [PDF]).
- Mohamed Gharbi, « Tiques traversant la Mare nostrum, quels risques ? », Bulletin de l'Académie Vétérinaire de France, no 2, (ISSN 2259-2385, DOI 10.4267/2042/70868, lire en ligne
 )
)
- Vourc'h G, Bridges VE, Gibbens J, De Groot BD, McIntyre L, Poland R, and Barnouin J 2006. Detecting emerging diseases in farm animals through clinical observations. Emerg. Infect. Dis.12 (2): 204-210.
- Hoogstraal H, Kaiser MN, Ormsbee RA, Osborn D.J, Helmy I & Gaber S (1967) Hyaloma (Hyalommina) Rhipicephaloides neumann (Ixodoidea. Ixodidae) : Its identiy, host, and ecology, and Rickettsia conori, R Prowazeki, and Coxiella burneti infections in rodent host in Egypt. Journal of medical entomology, 4(4), 391-400. (résumé).
- Kröber T, Guerin P. The Tick Blood Meal: From a Living Animal or from a Silicone Membrane? Altex 24 Special issue 2007. 39 - 41.
- Telford SR III, Wormser GP. Bartonella spp. transmission by ticks not established. Emerg Infect Dis. 2010 ; 16 (3) : 385 - 384. (Texte complet).
- Angelakis E, Billeter SA, Breitschwerdt EB, Chomel BB, Raoult D. Potential for tick-borne bartonelloses. Emerg Infect Dis. 2010 ; 16 (3) : 379 - 381. (Texte complet).
- Dantas-Torres F, Lorusso V, Testini G, de Paiva-Cavalcanti M, Figueredo LA, Stanneck D, Mencke N, Brandão-Filho SP, Alves LC, Otranto D. Detection of Leishmania infantum in Rhipicephalus sanguineus ticks from Brazil and Italy. Parasitol Res. 2010 Mar;106(4):857-60. (Résumé).
- Dantas-Torres F, Martins TF, de Paiva-Cavalcanti M, Figueredo LA, Lima BS, Brandão-Filho SP. Transovarial passage of Leishmania infantum kDNA in artificially infected Rhipicephalus sanguineus. Exp Parasitol. 2010 ;125 (2) :184-5. (Résumé).
- Paz GF, Ribeiro MF, Michalsky EM, da Rocha Lima AC, França-Silva JC, Barata RA, Fortes-Dias CL, Dias ES. Evaluation of the vectorial capacity of Rhipicephalus sanguineus (Acari: Ixodidae) in the transmission of canine visceral leishmaniasis. Parasitol Res. 2010 ; 106 (2) : 523-8. (Résumé).
- Walker DH, Dumler JS. Emergence of the Ehrlichioses as Human Health Problems. Emerging Infectious Diseases. 1996 ; 2 (1): 18-29 (Consulter ce texte en ligne). C'est une maladie contagieuse des ruminants et surtout des bovins, mais aussi des ovins et moindrement des caprins, due à une bactérie : Ehrlichia ruminantium autrefois nommé Cowdria ruminantium qui a donné son nom à cette maladie qui est donc aussi appelée ehrlichiose.
- Valls A, Pineda F, Belver M, Caballero T, López Serrano MC. Anaphylactic shock caused by tick (Rhipicephalus sanguineous). J Investig Allergol Clin Immunol. 2007 ; 17 (4) : 279-280. Texte complet, en anglais.
- Beaudouin E, Kanny G, Guerin B, Guerin L, Plenat F, Moneret- Vautrin DA. Unusual manifestations of hypersensitivity after a tick bite: report of two cases. Annals of Allergy, Asthma & Immunology. 1997;79:43-6.
- Moneret-Vautrin DA, Beaudouin E, Kanny G, Guerin L, Roche JF. Anaphylactic shock caused by ticks (Ixodes ricinus). J Allergy Clin Immunol. 1998;101:144-5.
- Gauci M, Loh R, Stone BF, Thong YH. Allergic reactions to the Australian paralysis tick, Ixodes holocyclus: diagnosis, evaluation by skin tests and radioimmunoassay. Clin Exp Allergy. 1989;19:279-83.
- Van Wye JE, Hsu Y, Lane R, Terr A, Moss R. Ig E antibodies in tick bite-induced anaphylaxis. J Allergy Clin Immunol. 1991;88(6):968-70.
- Acero S, Blanco R, Bartolomé B. Anaphylaxis due to a tick bite. Allergy. 2003;58(8):824-825.
- Domaine qui commence à être étudié en France en 2008, sur deux espèces principales (Ixodes ricinus et Dermacentor reticulatus) par l’équipe Génétique et Évolution des Interactions Hôte-Parasite du Laboratoire de biométrie et Biologie Évolutive (UMR CNRS 5558 Bat Mendel, 43 Bd 11 novembre 1918 - Villeurbanne), avec notamment Lionnel Zenner (École nationale vétérinaire de Lyon).
- Bouwknegt C, van Rijn PA, Schipper JJ, Hölzel D, Boonstra J, Nijhof AM, van Rooij EM, Jongejan F. Potential role of ticks as vectors of bluetongue virus. Exp Appl Acarol. 2010 Apr 1.
- Mellor PS (1990) The replication of bluetongue virus in Culicoides vectors. Curr Top Microbiol Immunol 162:143–161.
- Nevill EM. « Cattle and Culicoides biting midges as possible overwintering hosts of bluetongue virus. » Onderstepoort J Vet Res. 1971;38:65–72.
- A. Wilson, K. Darpel et P.S. Mellor, « Where does bluetongue virus sleep in the winter? », PLoS Biol., vol. 6, no 8, , p. 1612–1617 (DOI 10.1371/journal.pbio.0060210).
- J.L. Stott, B.I. Osburn et L. Alexander, « Ornithodoros coriaceus (pajaroello tick) as a vector of bluetongue virus », Am J Vet Res, vol. 46, , p. 1197–1199.
- Baumann D, Pusterla N, Peter O, Grimm F, Fournier PE, Schar G et al. Fieber nach Zeckenstich: Klinik und Diagnostik von akuten Zeckenstich-assoziierten Infektionskrankheiten in der Nordostschweiz. Dtsch Med Wochenschr. 2003; 128 (19) : 1042-7. (« Texte complet et résumé »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- Anke Hildebrandt, Jan Franke, Gernot Schmoock, Katharina Pauliks, Angela Krämer, and Eberhard Straube, Diversity and Coexistence of Tick-Borne Pathogens in Central Germany ; Journal of Medical Entomology 48(3):651-655. 2011 doi: 10.1603/ME10254 (Résumé, en anglais).
- Capek, I. (2006). Définition de priorités et actions réalisées dans le domaine des zoonoses non alimentaires, 2000-2005. Bulletin épidémiologique hebdomadaire 27-28, 196-199.
- Syndrome de mortalité des oiseaux lié aux tiques (« avian tick-related syndrome »), thèse de doctorat de Amélie, Charlotte PORCHE, École vétérinaire d'Alfort, PDF, 144 pages, 2011.
- MONKS D., FISHER M. et FORBES N.A. (2006) Ixodes frontalis and avian tick-related syndrome in the United Kingdom. Journal of small Animal Practice 47, 451-455.
- Beaudouin E, Thomas H, Nguyen-Grosjean V.M, Picaud J, Moumane L, Richard C, ... & Barbaud A (2015) Allergie à galactose-α1, 3 galactose (α-Gal) : une observation singulière et revue bibliographique. Revue Française d'Allergologie, 55(7), 492-497
- Lindgren E, Gustafson R. Tick-borne encephalitis in Sweden and climate change. The Lancet. 2001; 358:16-8. 9. ([travaux de l'Université de Stockholm).
- Communiqué Reuter du 9 juillet 2001 (consulté 2009 02 28).
- Sumilo, Asokliene, Bormane, Vasilenko, Golovljova & al., 2007 : Climate Change Cannot Explain the Upsurge of Tick-Borne Encephalitis in the Baltics. PLoS ONE, vol. 2, n. 6, e500 (texte intégral).
- Gritsun, TS; Nuttall, PA; Gould, E. Tick-borne flaviviruses. Advances in Virus research. 2003;61:317–371. PubMed.
- Charrel, RN; Attoui, H; Butenko, AM; Clegg, JC; Deubel, V, et al. Tick-borne virus diseases of human interest in Europe. Clinical microbiology and infection. 2004;10:1040–1055. [PubMed].
- A. Rizzoli, H.C. Hauffe, V. Tagliapietra, M. Neteler et R. Rosà, « Forest Structure and Roe Deer Abundance Predict Tick-Borne Encephalitis Risk in Italy », PLOS ONE, vol. 4, no 2, , e4336 (DOI 10.1371/journal.pone.0004336).
- P. Parola et al., « Warmer Weather Linked to Tick Attack and Emergence of Severe Rickettsioses », PLoS Neglected Tropical Diseases, vol. 2, no 11, , e338 (DOI 10.1371/journal.pntd.0000338).
- Jones K.E., Patel N.G., Levy M.A., Storeygard A., Balk D., Gittleman J.L. & Daszak P. 2008. Global trends in emerging infectious diseases. Nature, 451, 990-U4.
- Abbas, R. Z., Zaman, M. A., Colwell, D. D., Gilleard, J., & Iqbal, Z. (2014). Acaricide resistance in cattle ticks and approaches to its management: the state of play. Veterinary parasitology, 203(1-2), 6-20.
- Frisch JE. 1999. Towards a permanent solution for controlling cattle ticks. Int. J. Parasitol. 29: 57-71.
- Chevillon C. et al. 2007. Accumulation of acaricide resistance mechanisms in Rhipicephalus microplus (Acari: Ixodidae) populations from New Caledonia island. Vet. Parasitol. 147: 276-288.
- Ghosh S., Azhahianambi P. & Yadav M.P. 2007. Upcoming and future strategies of tick control: a review. Journal of Vector Borne Diseases, 44, 79-89.
- Pegram, RG, Tatchell RJ, de Castro JJ, Chizyuka HGB, Creek MJ, McCosker PJ, Moran MC & Nigarura G. 1993. Tick control: new concepts. World Anim. Rev. 74/75: 2-11.
- À propos de l'évaluation des facteurs biotiques et abiotiques d'émergence et de propagation de maladies transmises ou véhiculées par des tiques en Europe.
- Birulya NB, Zalutskaya TI. Warming of the climate and outbreaks of tick-borne encephalitis. Med Parazitol. (Mosk.) 1965 ; (3) : 65-67. (Langue : russe).
- J. Herchkovitch, « Maladie de Lyme : les renards à la rescousse : La maladie de Lyme a peut-être enfin rencontré sa némésis. La présence dans les forêts de renards réduirait le nombre de tiques infectées par la bactérie Borrelia burgdorferi. », sur Pourquoi Docteur, .
- (en) T.R. Hofmeester, P.A. Jansen, H.J Wijnen, E.C Coipan, M. Fonville, H.H.T. Prins, H. Sprong, et S.E. van Wieren, (2017) Cascading effects of predator activity on tick-borne disease risk | Proc. R. Soc. B 284: 20170453. |https://dx.doi.org/10.1098/rspb.2017.0453.
- Manches et jambes longues sont conseillées depuis longtemps par le personnel de l'ONF.
- (en) M. Brown et A.A. Hebert, Insect repellents: an overview, J Am Acad Dermatol, 1997;36:243-9.
- (en) M. Jensenius, A-M. Pretorius, F. Clarke, et B. Myrvang, Repellent efficacy of four commercial DEET lotions against Amblyomma hebraeum (Acari: Ixodidae), the principal vector of Rickettsia africae in southern Africa, Trans R Soc Trop Med Hyg, 2005;99:708-11.
- (en) J.K Trigg et N. Hill, Laboratory evaluation of a eucalyptus-based repellent against four biting arthropods, Phytother Res, 1996;10:313-6.
- (en) T.G.T. Jaenson, S. Garboui, et K. Pålsson, Repellency of oils of lemon eucalyptus, geranium, and lavender and the mosquito repellent MyggA Natural to Ixodes ricinus (Acari: Ixodidae) in the laboratory and field, J Med Entomol, 2006;43:731-6.
- (en) M.K. Faulde, W.M. Uedelhoven, M. Maleruis, et R.G Robbins, Factory-based Permethrin impregnation of uniforms: residual activity against Aedes aegypti and Ixodes ricinus in battle dress uniforms worn under field conditions, and cross-contamination during the laundering and storage process, Milit Med, 2006;171:472-7.
- (en) M.K. Faulde, W.M. Uedelhoven, et R.G Robbins, Contact toxicity and residual activity of different permethrin-based fabric impregnation methods for Aedes aegypti (Diptera: Culicidae), Ixodes ricinus (Acari: Ixodidae), and Lepisma saccharina (Thysanura: Lepismatidae), J Med Entomol, 2003;40:935-41.
- (en) C. Due, W. Fox, J.M. Medlock, M. Pietzsch, et L.G. Logan, Tick bite prevention and tick removal, BMJ, 2013;347:f7123.
- (en) Sood SK, Salzman MB, Johnson BJ, Happ CM, Piesman J et al., « Duration of tick attachment as a predictor of the risk of Lyme disease in an area in which Lyme disease is endemic », J Infect Dis, vol. 175, no 4, , p. 996-9. (PMID 9086168, lire en ligne [PDF]).
- (en) Gammons M, Salam G, « Tick removal », Am Fam Physician, vol. 66, no 4, , p. 643-5. (PMID 12201558, lire en ligne [html]).
- Gétaz L, Loutan L, Mezger N, « Maladies transmises par des tiques d’ici et d’ailleurs [Diseases transmitted by ticks locally and abroad] », Rev Med Suisse, vol. 8, no 340, , p. 974-6. (PMID 22662624, lire en ligne [html]).
- (en) Centers for Disease Control and Prevention, Tick removal, 2011.
- (en) De Boer R, Van den Bogaard AE, « Removal of attached nymphs and adults of Ixodes ricinus (Acari: Ixodidae) » J Med Entomol, 1993;30:748-52.
- (en) R.B. Nadelman, J. Nowakowski, D. Fish et al., « Prophylaxis with single-dose doxycycline for the prevention of Lyme disease after an Ixodes scapularis tick bite », N Engl J Med, vol. 345, , p. 79-84 (DOI 10.1056/NEJM200107123450201).
- J. Chmelar, E. Calvo, J.H.F. Pedra, I.M.B. Francischetti et M. Kotsyfakis, « Tick salivary secretion as a source of antihemostatics », Journal of Proteomics, vol. 75, no 13, , p. 3842-3854 (DOI 10.1016/j.jprot.2012.04.026, lire en ligne [PDF]).
- J.W.R. Hovius, M. Levi, E. Fikrig, « Salivating for knowledge: potential pharmacological agents in tick saliva », PLoS Med., vol. 5, no 2, , e43 (DOI 10.1371/journal.pmed.0050043).
- (en) L. Halos, T. Jamal, L. Vial, R. Maillard, A. Suau, A. Le Menach, HJ. Boulouis and M. Vayssier-Taussat, 2004a. Determination of an efficient and reliable method for DNA extraction from ticks. Vet Res. 5(35): 1-18.
- (en) L. Halos, T. Jamal, R. Maillard, F. Beugnet, A. Le Menach, HJ Boulouis and M. Vayssier-Taussat, 2005. Evidence of Bartonella sp. in questing adult and nymphal Ixodes ricinus ticks from France and co-infection with Borrelia burgdorferi sensu lato and Babesia sp. Vet. Res. 36: 79-87. FlecheH.
- Exemple : Collection de tiques de Montpellier ; incluant des Ixodidae et Argasidae, conservés dans de l'alcool pour la plupart. Ces tiques viennent essentiellement d'Afrique et du bassin méditerranéen, et moindrement d'Asie et d'Amérique. Des fiches sont disponibles pour chaque espèce, avec des montages entre lame et lamelle de larves, nymphes et organes adultes pour aider à la détermination.
- Tiques et maladies à tiques.
- (en) W.J. Tabachnick et W.C. Black IV, 1995. « Making a case for molecular population genetic studies of arthropod vectors ». Parasitol. Today 11: pp. 27-30.
- Action COST EurNegVec et projet EDENext.
- (en) M. Vayssier-Taussat, S. Moutailler, L. Michelet, E. Devillers, S. Bonnet, J. Cheval, et al. (2013) Next Generation Sequencing Uncovers Unexpected Bacterial Pathogens in Ticks in Western Europe. PLoS ONE 8(11): e81439. doi:10.1371/journal.pone.0081439
- (en) S. Bonnet, L. Michelet, S. Moutailler, J. Cheval, C. Hébert, M. Vayssier-Taussat, et M. Eloit, (2014), « Identification of parasitic communities within European ticks using next-generation sequencing ». PLoS Negl Trop Dis, 8(3), e2753.
- (en) M. Vayssier-Taussat, J. F. Cosson, B. Degeilh, M. Eloit, A. Fontanet, S. Moutailler, ... et P. Zylbermann (2015), « How a multidisciplinary ‘One Health' approach can combat the tick-borne pathogen threat in Europe ». Future Microbiology, 10(5), 809-818.
- (en) Diversity of viruses in Ixodes ricinus, and characterization of a neurotropic strain of Eyach virus.
- (en) S. Moutailler, I. Popovici, E. Devillers, M. Vayssier-Taussat, M. Eloit, New Microbes New Infect. 2016 Mar 5;11:71-81. doi: 10.1016/j.nmni.2016.02.012. eCollection, mai 2016.
- (en) Muriel Vayssier-Taussat, Sara Moutailler, Françoise Féménia, Philippe Raymond, Olivier Croce, Bernard La Scola, Pierre Edouard Fournier & Didier Raoult (2016), « Identification of Novel Zoonotic Activity of Bartonella spp. ». Emerging Infectious Disease Journal. March 2016, Volume 22, number 3
- (en) S. Moutailler, C. Valiente Moro, E. Vaumourin, L. Michelet, F.H. Tran, E. Devillers, et al. (2016), « Co-infection of Ticks: The Rule Rather Than the Exception ». PLoS Negl Trop Dis 10(3): e0004539. doi:10.1371/journal.pntd.0004539
- (en) C. Raileanu, S. Moutailler, I. Pavel, D. Porea, A.D. Mihalca, G. Savuta, et M. Vayssier-Taussat (2017), « Borrelia Diversity and Co-infection with Other Tick Borne Pathogens in Ticks ».Front Cell Infect Microbiol. 2017 Feb 14;7:36. doi: 10.3389/fcimb.2017.00036. eCollection.
- Tiques, maladie de Lyme et autres maladies à tiques, INRA, mai 2018, [PDF].
- CiTIQUE
- INRA
- CPIE
- ANSES
- DGS
- « Maladie de Lyme. Une application mobile pour lutter contre les tiques », Ouest-France, (consulté le ).
- signalement tique.
- Résultats intermédiaire publiés tous les deux mois par le projet CiTIQUE - https://www.citique.fr/les-chiffres-de-citique-juillet-aout-2017/
- CiTIQUE.
- Ohticks !.
- TéléTiQ.
- Nicot, Thresor de la langue française (1606).
- Dictionnaire de L'Académie française, 4e Édition (1762).
- Émile Littré, Dictionnaire de la langue française (1872-1877) ».
- Nouveau dictionnaire d'histoire naturelle, appliquée aux arts, à l'agriculture, à l'économie rurale et domestique, à la médecine etc, tome XXIX, Paris Cuez Deterville, libraire, rue Hautefeuille no 8 1819 (Consulter en ligne).
- Patrick Suskind, Le Parfum — Histoire d'un meurtrier, p. 26, traduction de l'allemand Bernard Lortholary, Paris, Fayard, 1985, réédition Librairie générale française, collection Le Livre de poche, 1988, 307 p. (ISBN 978-2-253-04490-1).
- Gilles Deleuze - L'Abécédaire de Gilles Deleuze avec Claire Parnet - Pierre-André Boutang (1996) - PART 1 - voir de 7 min 25 à 11 min 30.
- La boîte de Pandore et le fourre-tout de l'antiterrorisme espagnol, Non-fides, 18 décembre 2014.
Voir aussi
[modifier | modifier le code]Références taxonomiques
[modifier | modifier le code]- (en) Référence Tree of Life Web Project : Ixodida
- (en) Référence Catalogue of Life : Ixodida (consulté le )
- (en) Référence Fauna Europaea : Ixodida (consulté le )
- (en) Référence Paleobiology Database : Ixodida Leach 1815
- (fr + en) Référence ITIS : Ixodida
- (en) Référence NCBI : Ixodida (taxons inclus)
Bibliographie
[modifier | modifier le code]- Beati L & Klompen H (2017). Phylogeography of Ticks (Acari: Ixodida). Annual Review of Entomology, 63(1).
- Morel, Adam, Camicas & Hervy, 1998 : Les tiques du monde. Nomenclature, stades décrits, hôtes, répartition : The ticks of the world. Nomenclature, described stages, hosts, distribution (4 210 noms scientifiques, 869 espèces ou sous-espèces au ), /06/01 ; (ISBN 2-7099-1418-2) ; 233 pages.
- Nava, S., Venzal, J. M., Acuña, D. G., Martins, T. F., & Guglielmone, A. A. (2017). Ticks of the Southern Cone of America: Diagnosis, Distribution, and Hosts with Taxonomy, Ecology and Sanitary Importance. Academic Press |résumé).
- Pérez-Eid, 2007 : Les tiques. Identification, biologie, importance médicale et vétérinaire. Monographie de microbiologie, Tec & Doc EMinter- Lavoisier.
Articles connexes
[modifier | modifier le code]Liens externes
[modifier | modifier le code]- Attention aux tiques sur Service Public
- France Lyme, Association de lutte contre les Maladies Vectorielles à Tiques
- Enquête nationale sur la cartographie des morsures pour la France