
Radiodonta
| Radiodonta Temporal range:
| |
|---|---|

| |
| Left to right, top to bottom: Amplectobelua symbrachiata, Anomalocaris canadensis, Aegirocassis benmoulai, Peytoia nathorsti, Lyrarapax unguispinus, Cambroraster falcatus, and Hurdia victoria | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | †Radiodonta
Collins, 1996 |
| Families | |
| |
Radiodonta is an extinct order of stem-group arthropods that was successful worldwide during the Cambrian period. They may be referred to as radiodonts,[1][2][3] radiodontans,[4][5] radiodontids,[6] anomalocarids,[7] or anomalocaridids,[8][9][10] although the last two originally refer to the family Anomalocarididae, which previously included all species of this order but is now restricted to only a few species.[7] Radiodonts are distinguished by their distinctive frontal appendages, which are morphologically diverse and used for a variety of functions. Radiodonts included the earliest large predators known, but they also included sediment sifters and filter feeders.[11] Some of the most famous species of radiodonts are the Cambrian taxa Anomalocaris canadensis, Hurdia victoria, Peytoia nathorsti, Titanokorys gainessii, Cambroraster falcatus and Amplectobelua symbrachiata. The later surviving members include the subfamily Aegirocassisinae from the Early Ordovician of Morocco and the Early Devonian member Schinderhannes bartelsi from Germany.
Etymology
The name Radiodonta (Latin for radius "spoke of a wheel" and Greek for odoús "tooth") refers to the radial arrangement of tooth plates (oral cone) surrounding the mouth,[6] although these features are suggested to be absent in some radiodont species.[4][1]
Definition
The original diagnosis of order Radiodonta in 1996 is as follows:[6]
Radiodontids are bilaterally symmetrical, elongate arthropods with a nonmineralized cuticle typically most robust in the jaws and claws. The body is subdivided into two tagmata, much like the prosoma and opisthosoma of chelicerate arthropods. Typically, the front part shows no external segmentation, bears one pair of preoral claws, a pair of prominent eyes, and ventral jaws with radiating teeth. Some forms have additional rows of teeth and three or four postoral gnathobasic limb pairs. The trunk is metameric, typically with about 13 segments laterally developing imbricating lobes for swimming and gills for respiration, and may end in a prominent three-part tail. Some forms have gnathobasic trunk limbs.
In 2014, the clade Radiodonta was defined phylogenetically as a clade including any taxa closer to Anomalocaris canadensis than Paralithodes camtschaticus.[7] In 2019, it was redefined morphologically as animal bearing head carapace complex with central (H-) and lateral (P-) elements; outgrowths (endites) from frontal appendages bearing auxiliary spines; and reduced anterior flaps or bands of lamellae (setal blades) and strong tapering of body from anterior to posterior.[3]
Description

Most radiodonts were significantly larger than the other Cambrian fauna, with typical body lengths of large taxa varying from 30 to 50 cm (12 to 20 in).[12] The largest described radiodont is the Early Ordovician species Aegirocassis benmoulai, which may have grown up to 2 m (6.6 ft) long.[10][2] A nearly complete specimen of a juvenile Lyrarapax unguispinus measured only 18 mm (0.71 in), making it among the smallest radiodont specimens known, though adults reached a length of 8.3 cm (3.3 in)[2][13] An isolated frontal appendage of a hurdiid with a length less than half that of the juvenile Lyrarapax is known, but it is not known whether this specimen pertains to an adult.[14] The largest known Cambrian radiodont was Amplectobelua, reaching lengths of up to 90 cm (35 in) based on an incomplete specimen.[15] Anomalocaris canadensis was also relatively large, estimated up to 34.2–37.8 cm (13.5–14.9 in) long,[2] and the Cambrian hurdiid Titanokorys approached around 50 cm (20 in) long.[16]
The body of a radiodont could be divided into two regions: head and trunk. The head is composed of only one body segment[17] known as the ocular somite, covered by sclerites (head carapace complex), bore arthropodized frontal appendages, ventral mouthparts (oral cone), and stalked compound eyes. The tapering trunk is composed of multiple body segments, each associated with pairs of flaps and gill-like structures (setal blades).[3]
Frontal appendage


The anterior structures on the head are a pair of frontal appendages which have been referred to as 'claws', 'grasping appendages', 'feeding appendages', or 'great appendages' in previous studies (the last term is discouraged since the homology between frontal appendages and the original, morphologically distinct megacheiran great appendages is questionable.[17][18]). They are sclerotized (hardened) and arthropodized (segmented), bearing ventral endites (spines) on most of their podomeres (segmental units), and the endites may bear additional rows of auxiliary spines on their anterior and posterior margins.[19][3] The frontal appendage consists of two regions: the shaft ('peduncle',[2] 'base'[20] or 'promixal region'[2] in some studies) and the distal articulated region[19] (also referred to as 'claw'[20]). A triangular region covered by soft cuticle (arthrodial membrane) may occur on the ventral side between podomeres and provide flexibility.[21][11] Their purported pre-ocular and protocerebral origin suggest they are homologous to the primary antennae of Onychophora and the labrum of Euarthropoda (all arose from ocular somite),[17][9] while subsequent studies also suggest a deutocerebral origin and homologous with the chelicerae of Chelicerata and the antennae or 'great appendages' of other arthropods (all arose from post-ocular somite 1).[22] Since the morphology of the frontal appendages, especially those of the spines, always differs between species, it is one of the most important means of species identification.[19] In fact, many radiodonts are only known from a handful of fossilized frontal appendages.[21][19]
-
 Frontal appendages of Anomalocarididae, Amplectobeluidae, and possibly related species
Frontal appendages of Anomalocarididae, Amplectobeluidae, and possibly related species -
 Frontal appendages of Tamisiocarididae
Frontal appendages of Tamisiocarididae -
 Frontal appendages of Hurdiidae
Frontal appendages of Hurdiidae



Oral cone

The mouth is on the ventral side of the head, behind the attachment point of frontal appendages and is surrounded by a ring of tooth plates, forming the mouthpart known as oral cone ('jaws' in previous studies[6]). 3 or 4 tooth plates might be enlarged, giving the oral cone a triradial (e.g. Anomalocaris, Echidnacaris) or tetraradial (e.g. Hurdiidae, Lyrarapax) appearance.[23][13][24] The inner margin of tooth plates have spikes facing towards the mouth opening. Additional rows of internal tooth plates may occur in some hurdiid genera.[8][3] Detail reconstruction of some amplectobeluid oral cones are speculative, but they possibly did not present a typical radial arrangement.[4][1]
Head sclerites, eyes and trunk

Three head sclerite (carapace) complex formed by a central H-element (anterior sclerite or head shield) and a pair of P-elements (lateral sclerites) cover the dorsal and laterovental surface of the animal's head.[3] The P-elements may connect to each other as well as the H-element by a narrow anterior extension (P-element neck or 'beak').[8][3] The head sclerites are small and ovoid in Anomalocarididae and Amplectobeluidae,[4][3] but often enlarged in Hurdiidae, corresponded to their distinct body shapes (streamlined in Anomalocarididae/Amplectobeluidae but often compact in Hurdiidae).[3] The head bore two stalked compound eyes, which may have had mobility,[25] and are located between the gaps formed by the posterior regions of the H-element and P-elements.[8][3] The compound eyes of Echidnacaris are exceptionally unstalked.[14] Some species possess an additional median eye behind the H-element.[22]


Contrary to the original diagnosis, the division of body segments (segmental boundaries) can be visible externally[10][5][3] and no known member of Radiodonta (except the putative radiodont Cucumericrus[10][26]) is known to have pediform trunk appendages (legs).[27] The trunk has numerous body segments (somites), tapering from anterior to posterior, with the anterior three or four segments significantly constricted into a neck region.[3]
-
 Variations of radiodont body flaps
Variations of radiodont body flaps -
 The movement of radiodont flapping appendages
The movement of radiodont flapping appendages -
 Ventral view of a generalized GLS-bearing radiodont, showing gnathobase‐like structures (GLSs) associated with reduced anterior flaps
Ventral view of a generalized GLS-bearing radiodont, showing gnathobase‐like structures (GLSs) associated with reduced anterior flaps



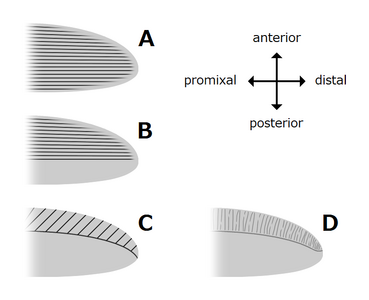
The trunk appendages were fin-like body flaps ('lateral flaps' or 'lobes' in some studies), usually one pair of ventral flaps per body segment, each slightly overlapping the one more anterior to it, but additional, non-overlapping sets of small dorsal flaps may occur in some Hurdiid species.[10] The flaps may have numerous vein-like structures (referred to as 'strengthening rays',[5] 'flap rays',[3] 'tranverse rods',[10] 'transverse lines'[28] or 'veins'[29]). The flaps on the neck region (referred to as 'reduced flaps',[4] 'neck flaps',[5] 'head flaps',[27] 'anterior flaps'[30] or 'differentiated flaps'[18]) are significantly reduced. In some species, jaw-like feeding appendages called gnathobase-like structures (GLSs) arose from each of the bases of their reduced neck flaps.[4][1] Numerous elongated blade-like extensions (referred to as lanceolate blades or lamellae[3]) arranged in a row, forming bands of gill-like structures known as setal blades, covered the dorsal surface of each body segment.[10] At least in Aegirocassis, each of the lanceolate blades are covered in wrinkles.[10] The ventral flaps may be homologous to the endopod of the biramous limbs of euarthropods and lobopodous limbs (lobopods) of gilled lobopodians, and the dorsal flaps and setal blades may be homologous to the exite and gill-bearing dorsal flaps of the former taxa.[31][10] The trunk may end either with a tail fan compose of 1 to 3 pairs of blades,[29][27][3] a pair of long furcae,[29][13][3] an elongated terminal structure,[27] or a featureless blunt tip.[10]
-
 Stanleycaris, Hurdia, Aegirocassis, Peytoia and Cambroraster are all examples of hurdiid radiodonts. These were the most diverse and long lasting of the radiodont families, surviving from the Cambrian up until the Devonian.
Stanleycaris, Hurdia, Aegirocassis, Peytoia and Cambroraster are all examples of hurdiid radiodonts. These were the most diverse and long lasting of the radiodont families, surviving from the Cambrian up until the Devonian. -
 Anomalocaris is a member of the anomalocarididae family, which at one point included all radiodonts, but now only includes a few genera such as Lenisicaris.
Anomalocaris is a member of the anomalocarididae family, which at one point included all radiodonts, but now only includes a few genera such as Lenisicaris. -
 Amplectobelua and Lyrarapax are representatives of the amplectobeluidae which is a very inclusive family of mainly Chinese radiodonts.
Amplectobelua and Lyrarapax are representatives of the amplectobeluidae which is a very inclusive family of mainly Chinese radiodonts. -
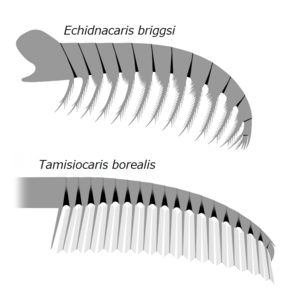
 Echidnacaris and Tamisiocaris are examples of the family tamisiocarididae which were exclusively suspension feeding radiodonts from the Cambrian.
Echidnacaris and Tamisiocaris are examples of the family tamisiocarididae which were exclusively suspension feeding radiodonts from the Cambrian.



Internal structures


Traces of muscles, digestive system and nervous system were described from some radiodont fossils. Pairs of well-developed muscles were connected to the ventral flaps located at the lateral cavities of each body segment.[27][9] Between the lateral muscles is a sophisticated digestive system, formed by a widening of the foregut and hindgut, both connected by a narrow midgut associated with six pairs of gut divercula (digestive glands).[27][5][32]
The brain of radiodonts was simpler than the three-segmented (compose of pro-, deuto- and tritocerebrum) brains of euarthropods, but further interpretations differ between studies. Based on Cong et al. 2014, the brain composed of only one brain segment originating from the ocular somite, the protocerebrum. The nerves of the frontal appendages and compound eyes arose from the anterior and lateral regions of the brain.[9][17] Based on Moysiuk & Caron 2022, the frontal appendage nerves arose from the ventral deutocerebrum, the second brain segment. The previous "frontal appendage nerves" actually represent median eye nerve.[22] In both interpretations, posterior to the brain was a pair of apparently unfused ventral nerve cords which ran through the animal's neck region.[9][22]
Paleoecology
Physiology

Radiodonts were interpreted as nektonic or nektobenthic animals, with their morphology suggesting an active swimming lifestyle. The muscular, overlapping ventral flaps may have propelled the animal through the water, possibly by moving in a wave-like formation resembling modern rays and cuttlefish.[33][34] Pairs of dorsal flaps, which make up a tail fan in some species, may have helped steering and/or stabilizing the animal during locomotion.[10][35] In Anomalocaris, morphology of the tail fan even suggests it could rapidly change its swimming direction efficiently.[36] On the other hand, some hurdiids have features significantly specialized for a nektobenthic lifestyle, such as Cambroraster with its dome-like H-element similar to the carapace of a horseshoe crab.[3] Bands of setal blades with wrinkling lanceolate blades may have increased the surface area, suggesting they were gills, providing the animal's respiratory function.[27][10] Abundance of the remains of scleritzed structures such as disarticulated frontal appendages and head sclerite complexes, suggest that mass moulting events may have occurred among radiodonts,[10][3] a behavior which also has been reported in some other Cambrian arthropods such as trilobites.[37]
Diet






Radiodonts had diverse feeding strategies, which could be categorized as raptorial predators, sediment sifters, or suspension, filter feeders.[2][38][11][39][40] For example, raptorial predators like Anomalocaris and Amplectobeluids might have been able to catch agile prey by using their raptorial frontal appendages; the latter even bore a robust endite for holding prey like a pincer.[26][21][4][11] With the smaller head carapace complex and large surface of arthrodial membranes, frontal appendages of these taxa had greater flexibility.[13] Stout frontal appendages of sediment sifters like Hurdia and Peytoia have serrated endites with mesial curvature, which could form a basket-like trap for raking through sediment and passing food items towards the well-developed oral cone.[3][11] Endites of frontal appendages from suspension/filter feeders like Tamisiocaris and Aegirocassis have flexible, densely-packed auxiliary spines, which could filter out organic components such as mesozooplankton and phytoplankton down to 0.5mm.[7][10] Frontal appendages of Caryosyntrips, which are unusual for radiodonts in having the direction of endite-bearing surfaces opposing one another and may have been able to manipulate and crush prey in a scissor-like slicing or grasping motion.[21][41]
Oral cones of radiodonts may have been used for suction and/or biting.[23][38][3] Together with the great variety of frontal appendages in different species of radiodonts, differentiation of oral cones between species suggests preferences of different diets as well.[38][11] For example, the triradial oral cone of Anomalocaris with irregular, tuberculated toothplates and a small opening may have been adapted to small and nektonic prey,[23][11] while the rigid tetraradial oral cones of Peytoia, Titanokorys, Hurdia, and one isolated oral cone attributed to Cambroraster with a larger opening and sometimes additional tooth plates may have been capable to consume larger food items relative to their body size and probably benthic or endobenthic prey.[23][38][3]
Classification
Taxonomic affinities
| |||||||||||||||||||||||||||||||||||||||||||||
| Summarized phylogeny between Radiodonta and other Ecdysozoan taxa[42] |





-

-
 The radiodont/opabiniid-euarthropod intermediate Kylinxia, shares many of the characteristics found in both dinocaridids and euarthropods
The radiodont/opabiniid-euarthropod intermediate Kylinxia, shares many of the characteristics found in both dinocaridids and euarthropods -
 A life reconstruction of the basal deuteropod Erratus, which helped reveal the evolution of arthropod trunk appendages
A life reconstruction of the basal deuteropod Erratus, which helped reveal the evolution of arthropod trunk appendages -
 Mieridduryn is a dinocaridid panarthropod from the Middle Ordovician that shares traits with both opabiniids and radiodonts
Mieridduryn is a dinocaridid panarthropod from the Middle Ordovician that shares traits with both opabiniids and radiodonts



Most phylogenetic analyses suggest that radiodonts, alongside opabiniids (Opabinia and Utaurora[43]), are stem-group arthropods just basal to deuteropoda,[42] a clade including upper stem (e.g. fuxianhuiids and bivalved arthropods) and crown Euarthropoda (e.g. Artiopoda, Chelicerata and Mandibulata).[8][44][45][46][47][48][7][9][10][2][3][30][18][39][40][49][50][43] This interpretation is supported by numerous arthropod groundplan found on radiodonts and opabiniids, such as stalked compound eyes,[25] digestive glands,[32] trunk appendages forming by dorsal and ventral elements (precursor of arthropod biramous appendages).[10][50] Compared to opabiniids, which possess posterior mouth opening and fused frontalmost appendages (comparable to euarthropod posterior-facing labrum/hypostome complex),[17][43] radiodonts on the other hand featured euarthropod-like dorsal sclerite (H-element) and arthropodization (frontal appendages) on their head regions,[51][17][43] alongside cuticularized gut termini.[27] The fact that both radiodonts and opabiniids lack exoskeleton on their trunk region suggests that the origin of compound eyes and arthropodization (segmented appendages) precede arthrodization (full set of trunk exoskeleton) in the arthropod stem lineage.[42][52][53] The constricted neck region with feeding appendicular structures of some radiodont may also shed light on the origin of the sophisticated arthropod head, which was formed by the fusion of multiple anterior body segments.[4][17] Basal deuteropods that possess a mixture of radiodont/opabiniid characters like Kylinxia and Erratus, may represent intermediate forms between radiodonts, opabiniids and other euarthropods.[18][50]
-
 The siberiid lobopodians Siberion (upper left), Megadictyon (bottom center) and Jianshanopodia (upper right)
The siberiid lobopodians Siberion (upper left), Megadictyon (bottom center) and Jianshanopodia (upper right) -

-
 Megacheira or 'great appendage arthropods', a class of possible stem-chelicerate previously thought to be radiodont's close relative
Megacheira or 'great appendage arthropods', a class of possible stem-chelicerate previously thought to be radiodont's close relative



Taxa just basal to the radiodont, opabiniid and euarthropod branch are 'gilled lobopodians' like Pambdelurion and Kerygmachela, which occasionally united under the class Dinocaridida with opabibiids and radiodonts.[54][46] They have body flaps, digestive glands, large (presumely compound) eyes and specialized frontal appendages like the former taxa, but their frontal appendages are not arthropodized nor fused, eyes sessile, gill-like structures less prominent, and certainlly bore lobopod underneath each of their flaps.[55][10][56][43] Taxa even basal to 'gilled lobopodians' are siberiids like Megadictyon and Jianshanopodia,[42] a group of lobopodians that bore robust frontal appendages and digestive glands, but no body flaps. Such intermediate forms between lobopodian and radiodont/euarthropod suggest that the total-group Arthropoda arose from a paraphyletic lobopodian grade, alongside the other two extant panarthropod phyla Tardigrada and Onychophora.[57][42][17][58][52][53]
Previous studies may suggest radiodonts as a group other than stem-arthropods, such as a hitherto unknown phylum;[33] cycloneuralian worms undergone convergent with arthropods (based on the cycloneuralian-like radial mouthparts);[59][54] stem chelicerate euarthropods alongside megacheirans also known as great appendage arthropods (based on the similarity between radiodont frontal appendages, megacheiran great appendages and chelicerae);[60] or Schinderhannes bartelsi, which resolved as a hurdiid radiodont in recent analyses,[42][7][10][2][3][39][40] as a species more closely related to euarthropods than other radiodonts (based on some putative euarthropod-like features found on Schinderhannes).[35] However, neither each of them were supported by later investigations. The radial mouthparts are not cycloneuralian-exclusive and more likely present result of convergent evolution or ecdysozoan plesimorphy, since they also have been found in panarthropods such as tardigrade and some lobopodians;[61] radiodonts lacking definitive euarthropod features such as trunk tergites and multiple head appendages,[42] and the megacheiran great appendages were considered to be deutocerebral,[62][63] which could be non-homologous to the radiodont protocerebral frontal appendages;[9][17] putative euarthropod characters found on the single Schinderhannes fossil is questionable and may present other radiodont-like structures.[42]
Interrelationships
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogeny of Radiodonta after Moysiuk & Caron 2021[39] |





Traditionally, all radiodont species have been placed within one family, Anomalocarididae,[6] hence the previous common name 'anomalocaridid'[26][8] and it was still occasionally used to refer the whole order even after reclassification.[9][10] Since the reassignment done by Vinther et al. 2014, most of the radiodont species were reclassified within three new families: Amplectobeluidae, Tamisiocarididae[2][3] (formerly Cetiocaridae[7]), and Hurdiidae.[7][10][2][3] Including Anomalocarididae, the four recent radiodont families may form the clade Anomalocarida.[7]
The original description of the order Radiodonta included Anomalocaris, Laggania (later known as Peytoia), Hurdia, Proboscicaris, Amplectobelua, Cucumericrus, and Parapeytoia.[6] However, Proboscicaris is now regarded as a junior synonym of Hurdia, and Parapeytoia is considered to be a Megacheiran.[8][27][10] Due to the limited discovery, The position of Cucumericrus within Radiodonta is unclear, as it was either unselected by phylogenetic analysis[7][3][2][39][40] or resolved in a polytomy with Radiodonta and Euarthropoda.[10][13]
-
 One of the poorly-known body parts (trunk appendage) of Cucumericrus decoratus; this species may not represent a true radiodont.
One of the poorly-known body parts (trunk appendage) of Cucumericrus decoratus; this species may not represent a true radiodont. -
 Frontal appendage of Echidnacaris briggsi, a tamisiocarid radiodont that was once suggested to belong to the Anomalocaris genus until its description in 2023.
Frontal appendage of Echidnacaris briggsi, a tamisiocarid radiodont that was once suggested to belong to the Anomalocaris genus until its description in 2023.

The first in-depth phylogenetic analysis of Radiodonta was conducted by Vinther et al. in 2014,[7] followed by a handful of subsequest studies with more or less modified results.[9][10][2][13][3][39][40][43] In most analysis, Caryosyntrips is the basal-most genus, but either resolved in a polytomy with other radiodonts and Euarthropoda (alongside Cucumericrus if included[10][13]) or outside of Radiodonta, casting doubt on its radiodont affinity.[64] With the exclusion of questionable Caryosyntrips and Cucumericrus, the monophyly of Radiodonta is widely supported,[7][9][10][2][13][3][39][40] with a few results suggest possible paraphyly (either the Anomalocarididae+Amplectobeluidae clade or Hurdiidae sister to Euarthropoda).[30][43] Putative synapomorphies of monophyletic Radiodonta including tripartite head sclerite complex and differentiated neck region.[3] The genus Anomalocaris in a broader sense always found to be polyphyletic, usually with "Anomalocaris" kunmingensis and "Anomalocaris" briggsi resolved as a member of Amplectobeluidae and Tamisiocarididae respectively.[7][9][10][2][3][39][40] Interrelationship of Amplectobeluidae is uncertain, as the amplectobeluid affinities of Lyrarapax and Ramskoeldia were occasionally questioned.[1][3][40] Monophyly of the speciose family Hurdiidae was recovered by most analysis and well-supported by several synapomorphies (e.g. distal articulated region of frontal appendage with proximal 5 podomeres bearing subequal endites[19][3]). Tamisiocarididae was often suggested to be sister group of Hurdiidae in 2010s,[7][10][2][3] but this position became questionable in subsequent studies.[22][24]
- Radiodonta
- ?Cucumericrus[26] (radiodont affinity questionable[65][64])
- ?Caryosyntrips[21] (radiodont affinity questionable[64])
- Anomalocarida
- Paranomalocaris[66] (placed within Anomalocarididae by some studies.[13][67])
- Laminacaris[68] (placed within Amplectobeluidae by some studies.[2])
- Houcaris (either placed within Anomalocarididae,[10][13][43] Amplectobeluidae[7][2] or Tamisiocarididae.[69][39])
- Innovatiocaris[70]
- Anomalocarididae
- Anomalocaris (in a broader sense, some species may placed within the other families.[7][10])
- Lenisicaris[20]
- Shucaris (either placed within Anomalocarididae or Amplectobeluidae[71][64])
- Amplectobeluidae
- Lyrarapax[9] (position questioned by some studies.[1])
- Amplectobelua[26]
- Ramskoeldia[1] (position questioned by some studies.[3])
- Guanshancaris[67][72]
- Shucaris (either placed within Anomalocarididae or Amplectobeluidae[71][64])
- Tamisiocarididae
- Hurdiidae/Peytoiidae[64]
- Aegirocassisinae[73]
- Peytoia
- Schinderhannes (position questioned by some studies.[74][75][76])
- Hurdia
- Stanleycaris[77]
- Pahvantia[2]
- Ursulinacaris[19]
- Cambroraster[3]
- ?Zhenghecaris (putative hurdiid radiodont[65])
- Cordaticaris[78]
- Buccaspinea[79]
- Titanokorys[16]
| Species | Original description | Year named | Family | Age | Location | Frontal appendage | Head sclerite complex |
|---|---|---|---|---|---|---|---|
| Cucumericrus decoratus | Hou, Bergström, & Ahlberg | 1995[26] | (unassigned) | Cambrian Stage 3 | Unknown | Unknown | |
| Caryosyntrips serratus | Daley & Budd | 2010[21] | (unassigned) | Wuliuan–Drumian | 
|
Unknown | |
| Caryosyntrips camurus | Pates & Daley | 2017[41] | (unassigned) | Wuliuan | 
|
Incomplete[79] | |
| Caryosyntrips durus | Pates & Daley | 2017[41] | (unassigned) | Drumian | 
|
Unknown | |
| Paranomalocaris multisegmentalis | Wang, Huang, & Hu | 2013[66] | Anomalocarididae? | Cambrian Stage 4 | 
|
Unknown | |
| Paranomalocaris simplex | Jiao, Pates, Lerosey-Aubril, Ortega-Hernandez, Yang, Lan, Zhang | 2021[67] | Anomalocarididae? | Cambrian Stage 4 | 
|
Unknown | |
| Laminacaris chimera | Guo, Pates, Cong, Daley, Edgecombe, Chen, & Hou | 2018[68] | (controversial) | Cambrian Stage 3 | 
|
Unknown | |
| Innovatiocaris maotianshanensis | Zeng, Zhao, Zhu | 2022[70] | (unassigned) | Cambrian Stage 3 | 
|
P-element unknown[70] | |
| Innovatiocaris? multispiniformis | Zeng, Zhao, Zhu | 2022[70] | (unassigned) | Cambrian Stage 3 | 
|
Unknown | |
| Anomalocaris canadensis | Whiteaves | 1892[80] | Anomalocarididae | Wuliuan | 
|

| |
| Lenisicaris pennsylvanica (formerly Anomalocaris pennsylvanica)[20] | Resser | 1929 | Anomalocarididae | Cambrian Stage 3 | 
|
Unknown | |
| Lenisicaris lupata | Wu, Ma, Lin, Sun, Zhang, & Fu | 2021[20] | Anomalocarididae | Cambrian Stage 3 | 
|
Unknown | |
| Anomalocaris daleyae | Paterson, García-Bellidob & Edgecombe | 2023 | Anomalocarididae | Cambrian Stage 4 | 
|
Unknown | |
| Houcaris magnabasis (formerly Anomalocaris magnabasis)[69] | Pates, Daley, Edgecombe, Cong & Lieberman | 2019 | (controversial) | Cambrian Stage 4 | 
|
Unknown | |
| Houcaris saron (formerly Anomalocaris saron)[69] | Hou, Bergström, & Ahlberg | 1995 | (controversial) | Cambrian Stage 3 | 
|
Unknown | |
| Echidnacaris briggsi[24] | Nedin | 1995 | Tamisiocarididae | Cambrian Stage 4 | 
|
Possible H-element and unique lateral sclerites associated with compound eyes[14][24] | |
| Ramskoeldia platyacantha | Cong, Edgecombe, Daley, Guo, Pates, & Hou | 2018[1] | Amplectobeluidae | Cambrian Stage 3 | 
|
Incomplete[1] | |
| Ramskoeldia consimilis | Cong, Edgecombe, Daley, Guo, Pates, & Hou | 2018[1] | Amplectobeluidae | Cambrian Stage 3 | 
|
Incomplete[1] | |
| Lyrarapax unguispinus | Cong, Ma, Hou, Edgecombe, & Strausfield | 2014[9] | Amplectobeluidae | Cambrian Stage 3 | 
|
P-element neck unknown | |
| Lyrarapax trilobus | Cong, Daley, Edgecombe, Hou, & Chen | 2016[5] | Amplectobeluidae | Cambrian Stage 3 | 
|
P-element unknown | |
| Amplectobelua symbrachiata | Hou, Bergström, & Ahlberg | 1995[26] | Amplectobeluidae | Cambrian Stage 3 | 
|

| |
| Amplectobelua stephenensis | Daley & Budd | 2010[21] | Amplectobeluidae | Wuliuan | 
|
Unknown | |
| Guanshancaris kunmingensis | Zhang et al. | 2023[72] | Amplectobeluidae | Cambrian Stage 4 | 
|
Unknown | |
| Tamisiocaris borealis | Daley & Peel | 2010 | Tamisiocarididae | Cambrian Stage 3 | 
|
Incomplete[7] | |
| Ursulinacaris grallae | Pates, Daley & Butterfield | 2019 | Hurdiidae | Wuliuan | 
|
Unknown | |
| Schinderhannes bartelsi | Kühl, Briggs, & Rust | 2009[35] | Hurdiidae | Emsian | Incomplete[3] | Incomplete[3] | |
| Stanleycaris hirpex | Pates, Daley, & Ortega-Hernández | 2018[77] | Hurdiidae | Wuliuan | 
|
P-element is unknown, possibly absent[22] | |
| Peytoia nathorsti | Walcott | 1911[81] | Hurdiidae | Wuliuan–Drumian | 
|
Incomplete[3] | |
| Peytoia infercambriensis (formerly Cassubia infercambriensis)[82] | Lendzion | 1975 | Hurdiidae | Cambrian Stage 3 | 
|
Unknown | |
| Aegirocassis benmoulai | Van Roy, Daley, & Briggs | 2015[10] | Hurdiidae (Aegirocassisinae) | Tremadocian | 
|

| |
| Hurdia victoria | Walcott | 1912[83] | Hurdiidae | Wuliuan–Drumian | 
|

| |
| Hurdia triangulata | Walcott | 1912[83] | Hurdiidae | Wuliuan |
|

| |
| Cambroraster falcatus | Moysiuk & Caron | 2019[3] | Hurdiidae | Wuliuan | 
|

| |
| Pahvantia hastata | Robison & Richards | 1981 | Hurdiidae | Drumian | 
|

| |
| Cordaticaris striatus | Sun, Zeng, & Zhao | 2020[78] | Hurdiidae | Drumian | Incomplete[78] | 
| |
| Zhenghecaris shankouensis | Vanner, Chen, Huang, Charbonnier, & Wang | 2006 | Hurdiidae | Cambrian Stage 3 | Unknown |  | |
| Buccaspinea cooperi | Pates, Lerosey-Aubril, Daley, Kier, Bonino & Ortega-Hernández | 2021[79] | Hurdiidae | Drumian | 
|
Unknown | |
| Titanokorys gainesi | Caron & Moysiuk | 2021[40] | Hurdiidae | Wuliuan | 
|

| |
| Pseudoangustidontus duplospineus | Van Roy & Tetlie | 2006 | Hurdiidae (Aegirocassisinae) | Tremadocian | 
|
Unknown | |
| Pseudoangustidontus izdigua | Potin, Gueriau & Daley | 2023 | Hurdiidae (Aegirocassisinae) | Tremadocian | 
|
Incomplete[73] | |
| Shucaris ankylosskelos | Wu et al. | 2024 | (controversial) | Cambrian Stage 2–Cambrian Stage 3 |  |
Incomplete[71] |
History

-
 Frontal appendage of Anomalocaris canadensis
Frontal appendage of Anomalocaris canadensis -
 Oral cone of Peytoia nathorsti
Oral cone of Peytoia nathorsti -
 H-element of Hurdia victoria
H-element of Hurdia victoria -
![Paired frontal appendages from an unnamed hurdiid radiodont[39]](//upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg)



The history of radiodonts is complex. Incomplete specimens pertaining to different body parts of the same species had historically been interpreted as belonging to different species and even different phyla.[6][8] Prior to their recognition as a group, radiodont specimens had been assigned to five different phyla: Porifera, Cnidaria, Echinodermata, Annelida, and Arthropoda.[6]
The first known radiodont specimens were collected from the trilobite beds of Mount Stephen by Richard G. McConnell of the Geological Survey of Canada in 1886[6] or 1888.[80] These specimens were named Anomalocaris canadensis in 1892 by GSC paleontologist Joseph Whiteaves.[80] Whiteaves interpreted the specimens, now known to be isolated frontal appendages, as the abdomen of a phyllocarid crustacean.[80] Additional radiodont specimens were described in 1911 by Charles Walcott.[81] He interpreted an isolated oral cone, which he named Peytoia nathorsti, as a jellyfish, and a poorly-preserved but relatively complete specimen, which he named Laggania cambria, as a holothurian.[81] In 1912 Walcott named Hurdia victoria and H. triangulata based on isolated H-elements, which he interpreted as the carapaces of crustaceans.[83] Isolated frontal appendages of Peytoia and Hurdia, collectively known as "Appendage F" in Briggs 1979, were all identified as those of Sidneyia at that time.[81] A Hurdia P-element was named Proboscicaris in 1962, and interpreted as the carapace of a bivalved arthropod.[84]
The Geological Survey of Canada initiated a revision of Burgess Shale fossils in 1966, overseen by Cambridge University paleontologist Harry B. Whittington.[6] This revision would ultimately lead to the discovery of the complete radiodont body plan. In 1978, Simon Conway Morris recognized that the mouthparts of Laggania were Peytoia-like, but he interpreted this as evidence that it was a composite fossil made up of a Peytoia jellyfish and a sponge.[85] In 1979, Derek Briggs recognized that the fossils of Anomalocaris were appendages, not abdomens, but interpreted them as walking legs alongside "Appendage F".[86] It was not until 1985 that the true nature of the fossils of Anomalocaris, Laggania, and Peytoia was recognized, and they were all assigned to a single genus, Anomalocaris.[33] Subsequently, it was recognized that Anomalocaris was a distinct form from the other two, resulting in a split into two genera, the latter of which was variously named Laggania and Peytoia until it was determined that both represent the same species and Peytoia had priority.[23] It was later recognized that some of the fossils assigned to these taxa belonged to another form, which was recognized as bearing a carapace made up of Hurdia and Proboscicaris elements. Finally, in 2009, these specimens were redescribed as Hurdia.[8] Even after these recognitions, partial misidentifications (e.g. oral cone and frontal appendages of Peytoia had been assigned to Anomalocaris[6] and Hurdia,[8] respectively) had been revealed by subsequent studies as well.[23][87]
The taxon Radiodonta itself was coined in 1996 by Desmond Collins, after it was established that Anomalocaris and its kin represented a distinctive lineage with arthropod affinities rather than a hitherto unknown phylum.[6] Collins also established the class Dinocarida to contain the order Radiodonta as well as the Opabiniidae, which he recognized as distinct due to its lacking the distinctive oral cone structure of radiodonts.[6] Radiodonta was first given a phylogenetic definition in 2014.[7] Radiodonta was originally viewed as containing a single family, Anomalocarididae, but it was divided into four families in 2014: Amplectobeluidae, Anomalocarididae, Cetiocaridae, and Hurdiidae.[7] The name Cetiocaridae did not conform to the International Code of Zoological Nomenclature and so was renamed Tamisiocarididae in 2019.[88]
Until the 2010s, radiodonts were typically considered to be uniformly large apex predators, but discoveries of new species over the course of that decade led to a considerable increase in the known ecological and morphological diversity of the group.[7][10][2][3][89][79][39][40]
References
- ^ a b c d e f g h i j k Cong, Pei-Yun; Edgecombe, Gregory D.; Daley, Allison C.; Guo, Jin; Pates, Stephen; Hou, Xian-Guang (2018). "New radiodonts with gnathobase-like structures from the Cambrian Chengjiang biota and implications for the systematics of Radiodonta". Papers in Palaeontology. 4 (4): 605–621. doi:10.1002/spp2.1219. ISSN 2056-2802. S2CID 90258934.
- ^ a b c d e f g h i j k l m n o p q r s t Lerosey-Aubril, Rudy; Pates, Stephen (14 September 2018). "New suspension-feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton". Nature Communications. 9 (1): 3774. Bibcode:2018NatCo...9.3774L. doi:10.1038/s41467-018-06229-7. ISSN 2041-1723. PMC 6138677. PMID 30218075. Dryad Data
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an Moysiuk, J.; Caron, J.-B. (14 August 2019). "A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources". Proceedings of the Royal Society B: Biological Sciences. 286 (1908): 20191079. doi:10.1098/rspb.2019.1079. PMC 6710600. PMID 31362637.
- ^ a b c d e f g h Cong, Peiyun; Daley, Allison C.; Edgecombe, Gregory D.; Hou, Xianguang (30 August 2017). "The functional head of the Cambrian radiodontan (stem-group Euarthropoda) Amplectobelua symbrachiata". BMC Evolutionary Biology. 17 (1): 208. doi:10.1186/s12862-017-1049-1. ISSN 1471-2148. PMC 5577670. PMID 28854872.
- ^ a b c d e f Cong, Peiyun; Daley, Allison C.; Edgecombe, Gregory D.; Hou, Xianguang; Chen, Ailin (September 2016). "Morphology of the radiodontan Lyrarapax from the early Cambrian Chengjiang biota". Journal of Paleontology. 90 (4): 663–671. doi:10.1017/jpa.2016.67. ISSN 0022-3360. S2CID 88742430.
- ^ a b c d e f g h i j k l m Collins, Desmond (1996). "The "evolution" of Anomalocaris and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.)". Journal of Paleontology. 70 (2): 280–293. doi:10.1017/S0022336000023362. S2CID 131622496.
- ^ a b c d e f g h i j k l m n o p q r s t Vinther, Jakob; Stein, Martin; Longrich, Nicholas R.; Harper, David A. T. (2014). "A suspension-feeding anomalocarid from the Early Cambrian" (PDF). Nature. 507 (7493): 496–499. Bibcode:2014Natur.507..496V. doi:10.1038/nature13010. PMID 24670770. S2CID 205237459.
- ^ a b c d e f g h i j Daley, Allison C.; Budd, Graham E.; Caron, Jean-Bernard; Edgecombe, Gregory D.; Collins, Desmond (2009). "The Burgess Shale anomalocaridid Hurdia and its significance for early euarthropod evolution". Science. 323 (5921): 1597–1600. Bibcode:2009Sci...323.1597D. doi:10.1126/science.1169514. PMID 19299617. S2CID 206517995.
- ^ a b c d e f g h i j k l m n Cong, Peiyun; Ma, Xiaoya; Hou, Xianguang; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (2014). "Brain structure resolves the segmental affinity of anomalocaridid appendages". Nature. 513 (7519): 538–42. Bibcode:2014Natur.513..538C. doi:10.1038/nature13486. PMID 25043032. S2CID 4451239.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2015). "Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps". Nature. 522 (7554): 77–80. Bibcode:2015Natur.522...77V. doi:10.1038/nature14256. PMID 25762145. S2CID 205242881.
- ^ a b c d e f g h De Vivo, Giacinto; Lautenschlager, Stephan; Vinther, Jakob (28 July 2021). "Three-dimensional modelling, disparity and ecology of the first Cambrian apex predators". Proceedings of the Royal Society B: Biological Sciences. 288 (1955): 20211176. doi:10.1098/rspb.2021.1176. PMC 8292756. PMID 34284622.
- ^ Foster, J. (2014). "Magical Mystery Tour: The Biological Psychedelia of the Burgess Shale". Cambrian Ocean World: Ancient Sea Life of North America. Indiana University Press. pp. 195–252. ISBN 978-0253011824.
- ^ a b c d e f g h i j Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A.; Shu, Degan; Paterson, John R. (1 November 2018). "Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan". National Science Review. 5 (6): 863–869. doi:10.1093/nsr/nwy057. ISSN 2095-5138.
- ^ a b c Pates, Stephen; Botting, Joseph P.; McCobb, Lucy M. E.; Muir, Lucy A. (2020). "A miniature Ordovician hurdiid from Wales demonstrates the adaptability of Radiodonta". Royal Society Open Science. 7 (6): 200459. Bibcode:2020RSOS....700459P. doi:10.1098/rsos.200459. ISSN 2054-5703. PMC 7353989. PMID 32742697.
- ^ Wu, Yu; Pates, Stephen; Pauly, Daniel; Zhang, Xingliang; Fu, Dongjing (3 November 2023). "Rapid growth in a large Cambrian apex predator". National Science Review. doi:10.1093/nsr/nwad284. ISSN 2095-5138. PMC 10833464.
- ^ a b Caron, J.-B.; Moysiuk, J. (September 2021). "A giant nektobenthic radiodont from the Burgess Shale and the significance of hurdiid carapace diversity". Royal Society Open Science. 8 (9): 210664. Bibcode:2021RSOS....810664C. doi:10.1098/rsos.210664. PMC 8424305. PMID 34527273.
- ^ a b c d e f g h i Ortega-Hernández, Javier; Janssen, Ralf; Budd, Graham E. (1 May 2017). "Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective". Arthropod Structure & Development. Evolution of Segmentation. 46 (3): 354–379. doi:10.1016/j.asd.2016.10.011. ISSN 1467-8039. PMID 27989966.
- ^ a b c d Zeng, Han; Zhao, Fangchen; Niu, Kecheng; Zhu, Maoyan; Huang, Diying (2020). "An early Cambrian euarthropod with radiodont-like raptorial appendages". Nature. 588 (7836): 101–105. Bibcode:2020Natur.588..101Z. doi:10.1038/s41586-020-2883-7. ISSN 1476-4687. PMID 33149303. S2CID 226248177.
- ^ a b c d e f Pates, Stephen; Daley, Allison C.; Butterfield, Nicholas J. (11 June 2019). "First report of paired ventral endites in a hurdiid radiodont". Zoological Letters. 5 (1): 18. doi:10.1186/s40851-019-0132-4. ISSN 2056-306X. PMC 6560863. PMID 31210962.
- ^ a b c d e Wu, Yu; Ma, Jiaxin; Lin, Weiliang; Sun, Ao; Zhang, Xingliang; Fu, Dongjing (1 May 2021). "New anomalocaridids (Panarthropoda: Radiodonta) from the lower Cambrian Chengjiang Lagerstätte: Biostratigraphic and paleobiogeographic implications". Palaeogeography, Palaeoclimatology, Palaeoecology. 569: 110333. Bibcode:2021PPP...569k0333W. doi:10.1016/j.palaeo.2021.110333. ISSN 0031-0182. S2CID 233565727.
- ^ a b c d e f g h Daley, Allison C.; Budd, Graham E. (2010). "New anomalocaridid appendages from the Burgess Shale, Canada". Palaeontology. 53 (4): 721–738. Bibcode:2010Palgy..53..721D. doi:10.1111/j.1475-4983.2010.00955.x. ISSN 1475-4983.
- ^ a b c d e f g Moysiuk, Joseph; Caron, Jean-Bernard (8 August 2022). "A three-eyed radiodont with fossilized neuroanatomy informs the origin of the arthropod head and segmentation". Current Biology. 32 (15): 3302–3316.e2. doi:10.1016/j.cub.2022.06.027. ISSN 0960-9822. PMID 35809569. S2CID 250361698.
- ^ a b c d e f Daley, Allison C.; Bergström, Jan (April 2012). "The oral cone of Anomalocaris is not a classic peytoia". Naturwissenschaften. 99 (6): 501–504. Bibcode:2012NW.....99..501D. doi:10.1007/s00114-012-0910-8. ISSN 0028-1042. PMID 22476406. S2CID 2042726.
- ^ a b c d Paterson, John R.; García-Bellido, Diego C.; Edgecombe, Gregory D. (1 January 2023). "The early Cambrian Emu Bay Shale radiodonts revisited: morphology and systematics". Journal of Systematic Palaeontology. 21 (1). doi:10.1080/14772019.2023.2225066. ISSN 1477-2019. S2CID 259719252.
- ^ a b Strausfeld, Nicholas J.; Ma, Xiaoya; Edgecombe, Gregory D.; Fortey, Richard A.; Land, Michael F.; Liu, Yu; Cong, Peiyun; Hou, Xianguang (August 2015). "Arthropod eyes: The early Cambrian fossil record and divergent evolution of visual systems". Arthropod Structure & Development. 45 (2): 152–172. doi:10.1016/j.asd.2015.07.005. PMID 26276096.
- ^ a b c d e f g Xian-Guang, Hou; Bergström, Jan; Ahlberg, Per (September 1995). "Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China". GFF. 117 (3): 163–183. doi:10.1080/11035899509546213. ISSN 1103-5897.
- ^ a b c d e f g h i Daley, Allison C.; Edgecombe, Gregory D. (2014). "Morphology of Anomalocaris canadensis from the Burgess Shale". Journal of Paleontology. 88 (1): 68–91. doi:10.1666/13-067. S2CID 86683798.
- ^ Daley, Allison C.; Paterson, John R.; Edgecombe, Gregory D.; García-Bellido, Diego C.; Jago, James B. (2013). "New anatomical information on Anomalocaris from the Cambrian Emu Bay Shale of South Australia and a reassessment of its inferred predatory habits". Palaeontology. 56 (5): 971–990. doi:10.1111/pala.12029. ISSN 1475-4983. S2CID 128955512.
- ^ a b c Chen, Jun-yuan; Ramsköld, Lars; Zhou, Gui-qing (27 May 1994). "Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators". Science. 264 (5163): 1304–1308. Bibcode:1994Sci...264.1304C. doi:10.1126/science.264.5163.1304. PMID 17780848. S2CID 1913482.
- ^ a b c Aria, Cédric; Zhao, Fangchen; Zeng, Han; Guo, Jin; Zhu, Maoyan (8 January 2020). "Fossils from South China redefine the ancestral euarthropod body plan". BMC Evolutionary Biology. 20 (1): 4. doi:10.1186/s12862-019-1560-7. ISSN 1471-2148. PMC 6950928. PMID 31914921.
- ^ Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2013). Anomalocaridids had two sets of lateral flaps. 57th Annual Meeting of The Paleontological Association. Zurich, Switzerland.
- ^ a b Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. (2 May 2014). "Sophisticated digestive systems in early arthropods". Nature Communications. 5 (1): 3641. Bibcode:2014NatCo...5.3641V. doi:10.1038/ncomms4641. ISSN 2041-1723. PMID 24785191.
- ^ a b c Whittington, Harry Blackmore; Briggs, Derek Ernest Gilmor (14 May 1985). "The largest Cambrian animal, Anomalocaris, Burgess Shale, British-Columbia". Philosophical Transactions of the Royal Society of London. B, Biological Sciences. 309 (1141): 569–609. Bibcode:1985RSPTB.309..569W. doi:10.1098/rstb.1985.0096.
- ^ Usami, Yoshiyuki (7 January 2006). "Theoretical study on the body form and swimming pattern of Anomalocaris based on hydrodynamic simulation". Journal of Theoretical Biology. 238 (1): 11–17. Bibcode:2006JThBi.238...11U. doi:10.1016/j.jtbi.2005.05.008. ISSN 0022-5193. PMID 16002096.
- ^ a b c Kühl, Gabriele; Briggs, Derek E. G.; Rust, Jes (6 February 2009). "A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany". Science. 323 (5915): 771–773. Bibcode:2009Sci...323..771K. doi:10.1126/science.1166586. ISSN 0036-8075. PMID 19197061. S2CID 47555807.
- ^ Sheppard, K. A.; Rival, D. E.; Caron, J.-B. (1 October 2018). "On the Hydrodynamics of Anomalocaris Tail Fins". Integrative and Comparative Biology. 58 (4): 703–711. doi:10.1093/icb/icy014. hdl:1974/22737. ISSN 1540-7063. PMID 29697774.
- ^ Daley, Allison; Drage, Harriet (1 September 2015). "The fossil record of ecdysis, and trends in the moulting behaviour of trilobites". Arthropod Structure & Development. 45 (2): 71–96. doi:10.1016/j.asd.2015.09.004. PMID 26431634.
- ^ a b c d De Vivo, Giacinto; Lautenschlager, Stephan; Vinther, Jakob (16 December 2016). "Reconstructing anomalocaridid feeding appendage dexterity sheds light on radiodontan ecology".
- ^ a b c d e f g h i j k Moysiuk, Joseph; Caron, Jean-Bernard (2021). "Exceptional multifunctionality in the feeding apparatus of a mid-Cambrian radiodont". Paleobiology. 47 (4): 704–724. Bibcode:2021Pbio...47..704M. doi:10.1017/pab.2021.19. ISSN 0094-8373. S2CID 236552819.
- ^ a b c d e f g h i j Caron, J.-B.; Moysiuk, J. (2021). "A giant nektobenthic radiodont from the Burgess Shale and the significance of hurdiid carapace diversity". Royal Society Open Science. 8 (9): 210664. Bibcode:2021RSOS....810664C. doi:10.1098/rsos.210664. PMC 8424305. PMID 34527273.
- ^ a b c Pates, S.; Daley, A. C. (2017). "Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada". Papers in Palaeontology. 3 (3): 461–470. doi:10.1002/spp2.1084. ISSN 2056-2802. S2CID 135026011.
- ^ a b c d e f g h Ortega-Hernández, Javier (December 2014). "Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848". Biological Reviews of the Cambridge Philosophical Society. 91 (1): 255–273. doi:10.1111/brv.12168. ISSN 1469-185X. PMID 25528950. S2CID 7751936.
- ^ a b c d e f g h Pates, Stephen; Wolfe, Joanna M.; Lerosey-Aubril, Rudy; Daley, Allison C.; Ortega-Hernández, Javier (9 February 2022). "New opabiniid diversifies the weirdest wonders of the euarthropod stem group". Proceedings of the Royal Society B: Biological Sciences. 289 (1968): 20212093. doi:10.1098/rspb.2021.2093. PMC 8826304. PMID 35135344.
- ^ Edgecombe, Gregory D. (1 March 2010). "Arthropod phylogeny: An overview from the perspectives of morphology, molecular data and the fossil record". Arthropod Structure & Development. Fossil Record and Phylogeny of the Arthropoda. 39 (2): 74–87. doi:10.1016/j.asd.2009.10.002. ISSN 1467-8039. PMID 19854297.
- ^ Legg, David; Sutton, Mark; Edgecombe, Gregory; Caron, Jean-Bernard (10 October 2012). "Cambrian bivalved arthropod reveals origin of arthrodization". Proceedings. Biological Sciences. 279 (1748): 4699–4704. doi:10.1098/rspb.2012.1958. PMC 3497099. PMID 23055069.
- ^ a b Legg, David (2013). "Multi-Segmented Arthropods from the Middle Cambrian of British Columbia (Canada)". Journal of Paleontology. 87 (3): 493–501. doi:10.1666/12-112.1. ISSN 0022-3360. S2CID 86725173.
- ^ Legg, David; Sutton, Mark; Edgecombe, Gregory (30 September 2013). "Arthropod fossil data increase congruence of morphological and molecular phylogenies". Nature Communications. 4: 2485. Bibcode:2013NatCo...4.2485L. doi:10.1038/ncomms3485. PMID 24077329.
- ^ Legg, David A.; Vannier, Jean (2013). "The affinities of the cosmopolitan arthropod Isoxys and its implications for the origin of arthropods". Lethaia. 46 (4): 540–550. doi:10.1111/let.12032. ISSN 1502-3931.
- ^ O’Flynn, Robert J.; Williams, Mark; Yu, Mengxiao; Harvey, Thomas H. P.; Liu, Yu (2022). "A new euarthropod with large frontal appendages from the early Cambrian Chengjiang biota". Palaeontologia Electronica. 25 (1): 1–21. doi:10.26879/1167. ISSN 1094-8074. S2CID 246779634.
- ^ a b c Fu, Dongjing; Legg, David A.; Daley, Allison C.; Budd, Graham E.; Wu, Yu; Zhang, Xingliang (28 March 2022). "The evolution of biramous appendages revealed by a carapace-bearing Cambrian arthropod". Philosophical Transactions of the Royal Society B: Biological Sciences. 377 (1847): 20210034. doi:10.1098/rstb.2021.0034. PMC 8819368. PMID 35125000. S2CID 246608509.
- ^ Ortega-Hernández, Javier (15 June 2015). "Homology of Head Sclerites in Burgess Shale Euarthropods". Current Biology. 25 (12): 1625–1631. doi:10.1016/j.cub.2015.04.034. ISSN 0960-9822. PMID 25959966.
- ^ a b Giribet, Gonzalo; Edgecombe, Gregory D. (17 June 2019). "The Phylogeny and Evolutionary History of Arthropods". Current Biology. 29 (12): R592–R602. doi:10.1016/j.cub.2019.04.057. ISSN 0960-9822. PMID 31211983. S2CID 189926344.
- ^ a b Edgecombe, Gregory D. (2 November 2020). "Arthropod Origins: Integrating Paleontological and Molecular Evidence". Annual Review of Ecology, Evolution, and Systematics. 51 (1): 1–25. doi:10.1146/annurev-ecolsys-011720-124437. ISSN 1543-592X. S2CID 225478171.
- ^ a b Xianguang, Hou; Jan, Jan Bergström; Jiayu, In Rong; Zongjie, Fang; Zhanghe, Zhou; Renbin, Zhan; Xiangdong, Wang; Xunlai, Yuan; Xianguang, Hou (2006), Dinocaridids – anomalous arthropods or arthropod-like worms?, CiteSeerX 10.1.1.693.5869
- ^ Budd, Graham E. (1998). "The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, Lower Cambrian, N Greenland)". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 89 (4): 249–290. doi:10.1017/S0263593300002418. ISSN 1473-7116. S2CID 85645934.
- ^ Young, Fletcher J.; Vinther, Jakob (2017). "Onychophoran-like myoanatomy of the Cambrian gilled lobopodian Pambdelurion whittingtoni". Palaeontology. 60 (1): 27–54. Bibcode:2017Palgy..60...27Y. doi:10.1111/pala.12269. hdl:1983/92180ef0-2205-4c65-9a70-90d59cfea2f4. ISSN 1475-4983. S2CID 55477207.
- ^ Edgecombe, Gregory D. (2010). "Arthropod phylogeny: An overview from the perspectives of morphology, molecular data and the fossil record". Arthropod Structure & Development. 39 (2–3): 74–87. doi:10.1016/j.asd.2009.10.002. ISSN 1467-8039. PMID 19854297.
- ^ Daley, Allison C.; Antcliffe, Jonathan B.; Drage, Harriet B.; Pates, Stephen (22 May 2018). "Early fossil record of Euarthropoda and the Cambrian Explosion". Proceedings of the National Academy of Sciences. 115 (21): 5323–5331. Bibcode:2018PNAS..115.5323D. doi:10.1073/pnas.1719962115. ISSN 0027-8424. PMC 6003487. PMID 29784780.
- ^ Xian-Guang, Hou; Bergström, Jan; Ahlberg, Per (September 1995). "Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China". GFF. 117 (3): 163–183. doi:10.1080/11035899509546213. ISSN 1103-5897.
- ^ Haug, Joachim T.; Waloszek, Dieter; Maas, Andreas; Liu, Yu; Haug, Carolin (March 2012). "Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian: MANTIS SHRIMP-LIKE CAMBRIAN PREDATORS". Palaeontology. 55 (2): 369–399. doi:10.1111/j.1475-4983.2011.01124.x. S2CID 82841481.
- ^ Smith, Martin R.; Caron, Jean-Bernard (June 2015). "Hallucigenia 's head and the pharyngeal armature of early ecdysozoans" (PDF). Nature. 523 (7558): 75–78. Bibcode:2015Natur.523...75S. doi:10.1038/nature14573. ISSN 1476-4687. PMID 26106857. S2CID 205244325.
- ^ Tanaka, Gengo; Hou, Xianguang; Ma, Xiaoya; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (October 2013). "Chelicerate neural ground pattern in a Cambrian great appendage arthropod". Nature. 502 (7471): 364–367. Bibcode:2013Natur.502..364T. doi:10.1038/nature12520. ISSN 1476-4687. PMID 24132294. S2CID 4456458.
- ^ Ortega-Hernández, Javier; Lerosey-Aubril, Rudy; Pates, Stephen (18 December 2019). "Proclivity of nervous system preservation in Cambrian Burgess Shale-type deposits". Proceedings of the Royal Society B: Biological Sciences. 286 (1917): 20192370. doi:10.1098/rspb.2019.2370. PMC 6939931. PMID 31822253.
- ^ a b c d e f McCall, Christian R. A. (13 December 2023). "A large pelagic lobopodian from the Cambrian Pioche Shale of Nevada". Journal of Paleontology: 1–16. doi:10.1017/jpa.2023.63. ISSN 0022-3360.
- ^ a b Zeng, Han; Zhao, Fangchen; Yin, Zongjun; Zhu, Maoyan (2 January 2018). "Morphology of diverse radiodontan head sclerites from the early Cambrian Chengjiang Lagerstätte, south-west China". Journal of Systematic Palaeontology. 16 (1): 1–37. doi:10.1080/14772019.2016.1263685. ISSN 1477-2019. S2CID 133549817.
- ^ a b Yuanyuan, Wang; Huang, Diying; Hu, Shixue (1 November 2013). "New anomalocardid frontal appendages from the Guanshan biota, eastern Yunnan". Chinese Science Bulletin. 58 (32): 3937–3942. Bibcode:2013ChSBu..58.3937W. doi:10.1007/s11434-013-5908-x. S2CID 83910779.
- ^ a b c Jiao, De-guang; Pates, Stephen; Lerosey-Aubril, Rudy; Ortega-Hernandez, Javier; Yang, Jie; Lan, Tian; Zhang, Xi-guang (2021). "The endemic radiodonts of the Cambrian Stage 4 Guanshan biota of South China". Acta Palaeontologica Polonica. 66. doi:10.4202/app.00870.2020. ISSN 0567-7920. S2CID 236683029.
- ^ a b Guo, J.; Pates, S.; Cong, P.; Daley, A. C.; Edgecombe, G. D.; Chen, T.; Hou, X. (2018). "A new radiodont (stem Euarthropoda) frontal appendage with a mosaic of characters from the Cambrian (Series 2 Stage 3) Chengjiang biota". Papers in Palaeontology. 5 (1). ISSN 2056-2799.
- ^ a b c Wu, Yu; Fu, Dongjing; Ma, Jiaxin; Lin, Weiliang; Sun, Ao; Zhang, Xingliang (1 June 2021). "Houcaris gen. nov. from the early Cambrian (Stage 3) Chengjiang Lagerstätte expanded the palaeogeographical distribution of tamisiocaridids (Panarthropoda: Radiodonta)". PalZ. 95 (2): 209–221. doi:10.1007/s12542-020-00545-4. ISSN 1867-6812. S2CID 235221043.
- ^ a b c d Zeng, Han; Zhao, Fangchen; Zhu, Maoyan (7 September 2022). "Innovatiocaris, a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta". Journal of the Geological Society. 180. doi:10.1144/jgs2021-164. ISSN 0016-7649. S2CID 252147346.
- ^ a b c Wu, Yu; Pates, Stephen; Liu, Cong; Zhang, Mingjing; Lin, Weiliang; Ma, Jiaxin; Wu, Yuheng; Chai, Shu; Zhang, Xingliang; Fu, Dongjing (31 December 2024). "A new radiodont from the lower Cambrian (Series 2 Stage 3) Chengjiang Lagerstätte, South China informs the evolution of feeding structures in radiodonts". Journal of Systematic Palaeontology. 22 (1). doi:10.1080/14772019.2024.2364887. ISSN 1477-2019.
- ^ a b Zhang, Mingjing; Wu, Yu; Lin, Weiliang; Ma, Jiaxin; Wu, Yuheng; Fu, Dongjing (11 April 2023). "Amplectobeluid Radiodont Guanshancaris gen. nov. from the Lower Cambrian (Stage 4) Guanshan Lagerstätte of South China: Biostratigraphic and Paleobiogeographic Implications". Biology. 12 (4): 583. doi:10.3390/biology12040583. ISSN 2079-7737. PMC 10136193. PMID 37106783.
- ^ a b Potin, Gaëtan J.-M.; Gueriau, Pierre; Daley, Allison C. (2023). "Radiodont frontal appendages from the Fezouata Biota (Morocco) reveal high diversity and ecological adaptations to suspension-feeding during the Early Ordovician". Frontiers in Ecology and Evolution. 11. doi:10.3389/fevo.2023.1214109. ISSN 2296-701X.
- ^ Ortega-Hernández, Javier (December 2014). "Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848". Biological Reviews of the Cambridge Philosophical Society. 91 (1): 255–273. doi:10.1111/brv.12168. ISSN 1469-185X. PMID 25528950. S2CID 7751936.
- ^ Zhu, X.; Lerosey-Aubril, R.; Ortega-Hernández, J. (2021). "Furongian (Jiangshanian) occurrences of radiodonts in Poland and South China and the fossil record of the Hurdiidae". PeerJ. 9. e11800. doi:10.7717/peerj.11800. PMC 8312493. PMID 34386302.
- ^ Potin, Gaëtan J.-M.; Daley, Allison C. (2023). "The significance of Anomalocaris and other Radiodonta for understanding paleoecology and evolution during the Cambrian explosion". Frontiers in Earth Science. 11. Bibcode:2023FrEaS..1160285P. doi:10.3389/feart.2023.1160285. ISSN 2296-6463.
- ^ a b Pates, Stephen; Daley, Allison; Ortega-Hernández, Javier (3 March 2018). Reply to Comment on "Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris" with the formal description of Stanleycaris.
- ^ a b c Sun, Zhixin; Zeng, Han; Zhao, Fangchen (1 August 2020). "A new middle Cambrian radiodont from North China: Implications for morphological disparity and spatial distribution of hurdiids". Palaeogeography, Palaeoclimatology, Palaeoecology. 558: 109947. Bibcode:2020PPP...558j9947S. doi:10.1016/j.palaeo.2020.109947. ISSN 0031-0182. S2CID 224868404.
- ^ a b c d Pates, Stephen; Lerosey-Aubril, Rudy; Daley, Allison C.; Kier, Carlo; Bonino, Enrico; Ortega-Hernández, Javier (19 January 2021). "The diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian: Drumian)". PeerJ. 9: e10509. doi:10.7717/peerj.10509. ISSN 2167-8359. PMC 7821760. PMID 33552709.
- ^ a b c d Whiteaves, J. F. (1892). "Description of a new genus and species of phyllocarid Crustacea from the Middle Cambrian of Mount Stephen, B. C.". The Canadian Record of Science. 5 (4).
- ^ a b c d Walcott, C. D. 1911a. Middle Cambrian holothurians and medusae. Cambrian geoogy and paleontology II. Smithsonian Miscellaneous Collections, 57: 41–68.
- ^ Daley, Allison; Legg, David (2 July 2015). "A morphological and taxonomic appraisal of the oldest anomalocaridid from the Lower Cambrian of Poland". Geological Magazine. -1 (5): 949–955. Bibcode:2015GeoM..152..949D. doi:10.1017/S0016756815000412. S2CID 130745134.
- ^ a b c WALCOTT, C. D. 1912. Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata. Smithsonian Miscellaneous Collections, 57: 145–228.
- ^ ROLFE, W. D. I. 1962. Two new arthropod carapaces from the Burgess Shale (Middle Cambrian) of Canada. Breviora Museum of Comparative Zoology, 60: 1–9.
- ^ Conway Morris, S. (1978). "Laggania cambria Walcott: A Composite Fossil". Journal of Paleontology. 52 (1): 126–131. JSTOR 1303799.
- ^ Briggs, D. E. G. (1979). "Anomalocaris, the largest known Cambrian arthropod". Palaeontology. 22 (3): 631–664.
- ^ Daley, Allison; Budd, Graham; Caron, Jean-Bernard (1 October 2013). "The morphology and systematics of the anomalocarid Hurdia from the Middle Cambrian of British Columbia and Utah". Journal of Systematic Palaeontology. 11. doi:10.1080/14772019.2012.732723. S2CID 86465719.
- ^ Pates, Stephen; Daley, Allison C. (2019). "The Kinzers Formation (Pennsylvania, USA): the most diverse assemblage of Cambrian Stage 4 radiodonts". Geological Magazine. 156 (7): 1233–1246. Bibcode:2019GeoM..156.1233P. doi:10.1017/S0016756818000547. S2CID 134299859.
- ^ Paterson, John R.; Edgecombe, Gregory D.; García-Bellido, Diego C. (2020). "Disparate compound eyes of Cambrian radiodonts reveal their developmental growth mode and diverse visual ecology". Science Advances. 6 (49). Bibcode:2020SciA....6.6721P. doi:10.1126/sciadv.abc6721. hdl:10141/622906. PMC 7821881. PMID 33268353.
| ||||||||||||||||||
| ||||||||||||||||||